



Carbon isotope discrimination in C3 land plants is independent of natural variations in pCO2
Affiliations | Corresponding Author | Cite asKohn, M.J. (2016) Carbon isotope discrimination in C3 land plants is independent of natural variations in pCO2. Geochem. Persp. Let. 2, 35-43.
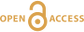

- Share this article





-
Article views:14,048Cumulative count of HTML views and PDF downloads.
- Download Citation
- Rights & Permissions

Abstract
 =
=Figures and Tables
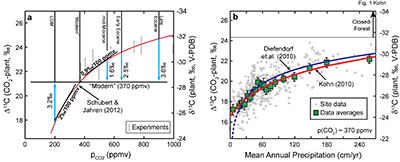 Figure 1 Proposed models for factors that influence δ13C of C3 plants. (a) pCO2. Differences are illustrated between geological conditions vs. AD 2000 (pCO2 = 370 ppmv, average δ13C = -28.5 for C3 biomass). LGM = Last Glacial Maximum. Note inverse relationship between δ13C and Δ13C. Experiments are for above-ground biomass (Schubert and Jahren, 2012), shifted to fit preferred curve. (b) Mean annual precipitation (data and data averages from Kohn, 2010). | 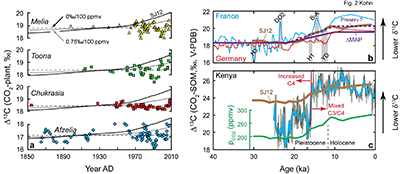 Figure 2 Tree ring and Pleistocene-Holocene records. (a) Tropical data from Thailand show near-zero pCO2 effect. Models are anchored on oldest 5-10 analyses (where any pCO2 effects should be smallest). SJ12 = model of Schubert and Jahren (2012). (b) Correction for changes in mean annual precipitation (“ΔMAP”), and possibly for organic matter preservation (“preserv.”) reproduces records independent of a direct pCO2-dependence. Brief climatic events also correlate with changes in Δ13C independent of pCO2. H1, H3 = Heinrich events 1 and 3; DO2 = Dansgaard-Oeschger event 2; B-A = Bølling-Allerød; YD = Younger Dryas. (c) An abrupt shift at c. 15.8 ka could reflect an abrupt threshold response of C4 biomass in the local ecosystem or variable contributions of diatoms to SOM (Fig. S-1). |
| Figure 1 | Figure 2 |
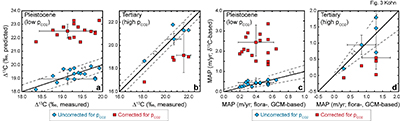 Figure 3 Cenozoic herbivore data. (a-d) Calculated vs. measured Δ13C, and calculated vs. independently estimated MAP, showing good correspondence with pCO2-independent model. Representative error bars (2σ) reflect propagated uncertainties in isotope compositions and tissue-diet fractionations, the dependence of δ13C on MAP, and proxy estimates of MAP and pCO2 (Table S-2). | 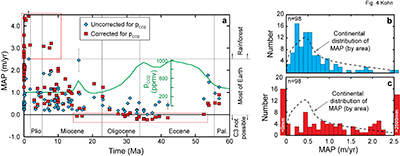 Figure 4 (a) Estimates of MAP for the Cenozoic based on fossil herbivore isotope compositions show a U-shaped distribution. pCO2-corrections imply widespread rainforests in the Pleistocene, and hyper-deserts in mid-Cenozoic. pCO2 curve simplified from Beerling and Royer (2011). (b-c) Histograms of Cenozoic MAP estimates show a better match between the global distribution (by area) of MAP (Kohn, 2010) vs. MAP estimated using the pCO2-independent model. The pCO2-dependent model predicts unusually high proportions of deserts (MAP < 0.1 m/yr) and rainforests (MAP > 2.5 m/yr) in the geologic record. |
| Figure 3 | Figure 4 |
Supplementary Figures and Tables
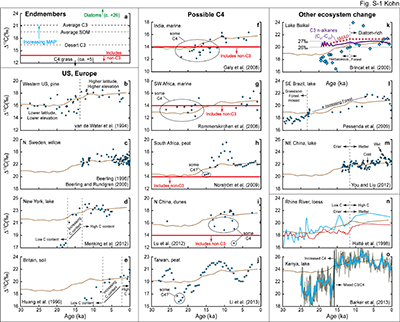 Figure S-1 (a) Principles of carbon isotope compositions relevant to interpreting isotopic records of sediment organic matter (SOM). C4 grasses have extremely low Δ13C, C3 plants have increasing Δ13C with increasing MAP, and diatoms have extremely high Δ13C. Applying a ~2 ‰ offset between plants and SOM (Bowling et al., 2008) implies a minimum cutoff of Δ13C(CO2-SOM) ~ 14 ‰ for which other sources of carbon besides C3 plants must be considered. (b-o) Records of Pleistocene-Holocene transition annotated with observations and interpretations from the original publications. |  Table S-1 Specific regression results for leaf and tree ring δ13C. | Table S-2 Herbivore isotope data and calculations of Δ13C and MAP. | Table S-3 Calculated pCO2-dependencies of δ13C in C3 plants. | Table S-4 Summary of records and factors influencing calculations. |
| Figure S-1 | Table S-1 | Table S-2 | Table S-3 | Table S-4 |
top
Introduction
C3 plants (trees, shrubs, herbs and cool-climate grasses) constitute c. 95 % of terrestrial plant biomass (Still et al., 2003
Still, C.J., Berry, J.A., Collatz, G.J., DeFries, R.S. (2003) Global distributions of C3 and C4 vegetation: carbon cycle implications. Global Biogeochemical Cycles 17, doi: 10.1029/2001GB001807.
), so understanding C3 plant response to changes in the partial pressure of atmospheric CO2 (pCO2) is key for modelling Earth’s carbon cycle (e.g., Cao et al., 2010Cao, L., Bala, G., Caldeira, K., Nemani, R., Ban-Weiss, G. (2010) Importance of carbon dioxide physiological forcing to future climate change. Proceedings of the National Academy of Sciences 107, 9513-9518, doi:10.1073/pnas.0913000107.
; Shevliakova et al., 2013Shevliakova, E., Stouffer, R.J., Malyshev, S., Krasting, J.P., Hurtt, G.C., Pacala, S.W. (2013) Historical warming reduced due to enhanced land carbon uptake. Proceedings of the National Academy of Sciences 110, 16730-16735, doi: 10.1073/pnas.1314047110.
). Carbon isotope compositions (δ13C) of C3 plants are a principal means of inferring water use efficiency (Farquhar et al., 1989Farquhar, G.D., Ehleringer, J.R., Hubick, K.T. (1989) Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 40, 503-537.
), which is important to models of future C3 biomass, and have been proposed to allow estimation of mean annual precipitation (MAP) in the geologic past (Kohn, 2010Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
). Plant δ13C depends on the δ13C of atmospheric CO2 (Freyer and Wiesberg, 1973Freyer, H.D., Wiesberg, L. (1973) 13C decrease in modern wood due to large-scale combustion of fossil fuels. Naturwissenschaften 60, 517-518.
), but an unresolved decades-long debate centres on whether terrestrial C3 plant δ13C responds to pCO2. Many studies have argued that increasing pCO2 increases carbon isotope discrimination (e.g., Feng and Epstein, 1995Feng, X., Epstein, S. (1995) Carbon isotopes of trees from arid environments and implications for reconstructing atmospheric CO2 concentration. Geochimica et Cosmochimica Acta 59, 2599-2608.
; Schubert and Jahren, 2012Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
); others have argued for no pCO2-effect (e.g., Arens et al., 2000Arens, N.C., Jahren, A.H., Amundson, R.G. (2000) Can C3 plants faithfully record the carbon isotopic composition of atmospheric carbon dioxide? Paleobiology 26, 137-164.
; Saurer et al., 2004Saurer, M., Siegwolf, R.T.W., Schweingruber, F.H. (2004) Carbon isotope discrimination indicates improving water-use efficiency of trees in northern Eurasia over the last 100 years. Global Change Biology 10, 2109-2120.
; Frank et al., 2015Frank, D.C., Poulter, B., Saurer, M., Esper, J., Huntingford, C., Helle, G., Treydte, K., Zimmermann, N.E., Schleser, G.H., Ahlström, A., Ciais, P., Friedlingstein, P., Levis, S., Lomas, M., Sitch, S., Viovy, N., Andreu-Hayles, L., Bednarz, Z., Berninger, F., Boettger, T., D‘Alessandro, C.M., Daux, V., Filot, M., Grabner, M., Gutierrez, E., Haupt, M., Hilasvuori, E., Jungner, H., Kalela-Brundin, M., Krapiec, M., Leuenberger, M., Loader, N.J., Marah, H., Masson-Delmotte, V., Pazdur, A., Pawelczyk, S., Pierre, M., Planells, O., Pukiene, R., Reynolds-Henne, C.E., Rinne, K.T., Saracino, A., Sonninen, E., Stievenard, M., Switsur, V.R., Szczepanek, M., Szychowska-Krapiec, E., Todaro, L., Waterhouse, J.S., Weig, M. (2015) Water-use efficiency and transpiration across European forests during the Anthropocene. Nature Climate Change 5, 579–583, doi: 10.1038/NCLIMATE2614.
). Recent work (Schubert and Jahren, 2012Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
) integrated natural and experimental data to infer that C3 plant δ13C depends hyperbolically on pCO2 (Fig. 1a) and that changes to carbon isotope compositions in leaves and sediment organic matter over the last 30 ka parallel and predict pCO2 (Schubert and Jahren, 2015Schubert, B.A., Jahren, A.H. (2015) Global increase in plant carbon isotope fractionation following the Last Glacial Maximum caused by increase in atmospheric pCO2. Geology 43, 435-438.
). In this study, carbon isotope records ranging from the last several decades through the early Cenozoic are critically evaluated to identify which records indeed provide quantitative constraints on pCO2-dependencies, and to provide a robust estimate of that dependence.Multiple factors besides atmospheric CO2 can affect C3 plant δ13C, including MAP, ontogeny, light levels, genetics and fertilisation (e.g., through atmospheric nitrogen deposition; Farquhar et al., 1989
Farquhar, G.D., Ehleringer, J.R., Hubick, K.T. (1989) Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 40, 503-537.
; Stewart et al., 1995Stewart, G.R., Turnbull, M.H., Schmidt, S., Erskine, P.D. (1995) 13C natural abundance in plant communities along a rainfall gradient: a biological integrator of water availability. Australian Journal of Plant Physiology 22, 51-55.
; McCarroll and Loader, 2004McCarroll, D., Loader, N.J. (2004) Stable isotopes in tree rings. Quaternary Science Reviews 23, 771-801.
; Diefendorf et al., 2010Diefendorf, A.F., Mueller, K.E., Wing, S.L., Koch, P.L., Freeman, K.H. (2010) Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proceedings of the National Academy of Sciences 107, 5738-5743.
; Kohn, 2010Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
; Fig. 1b). Other physical factors, including latitude and altitude, have a minor impact on C3 plant δ13C (Diefendorf et al., 2010Diefendorf, A.F., Mueller, K.E., Wing, S.L., Koch, P.L., Freeman, K.H. (2010) Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proceedings of the National Academy of Sciences 107, 5738-5743.
; Kohn, 2010Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
), but high variability both within and among trees must reflect individual responses to other, less easily identified, environmental or physiological factors (McCarroll and Loader, 2004McCarroll, D., Loader, N.J. (2004) Stable isotopes in tree rings. Quaternary Science Reviews 23, 771-801.
). Thus, records of natural δ13C through time, e.g., from tree rings, sediment records, etc. must account for changes to MAP and other environmental changes, most recently from anthropogenic activities, before any pCO2-dependence can be quantified.“Modern” conditions anchor many quantitative estimates of palaeoclimate, e.g., δ13C-based estimates of MAP are normalised to AD 2000 data (pCO2 = 370 ppmv, δ13CCO2 = ‑8.0 ‰). If C3 plant δ13C does depend on pCO2, δ13C values from periods of low pCO2 (e.g., late Pleistocene) are anomalously high, and must be downward corrected for any calculations (Δ13C increases); conversely δ13C values from periods of high pCO2 (e.g., Eocene) must be upward corrected (Δ13C decreases; Fig. 1a). In comparison with the modern dependence of C3 plant δ13C on MAP (Fig. 1b), uncorrected estimates of MAP must be too dry for the Pleistocene and too wet for the Eocene. Fortunately, estimates of MAP for many times are known from independent proxies (e.g., floral analysis) and general circulation models, and pCO2 is known either from measurements in ice cores or from geochemical proxies. Thus, pCO2-effects on C3 δ13C values can now be evaluated from ancient sediments and fossils, albeit with careful consideration of sometimes substantial errors in proxy estimates of pCO2 and MAP (see Supplementary Information).
Figure 1 Proposed models for factors that influence δ13C of C3 plants. (a) pCO2. Differences are illustrated between geological conditions vs. AD 2000 (pCO2 = 370 ppmv, average δ13C = -28.5 for C3 biomass). LGM = Last Glacial Maximum. Note inverse relationship between δ13C and Δ13C. Experiments are for above-ground biomass (Schubert and Jahren, 2012
Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
), shifted to fit preferred curve. (b) Mean annual precipitation (data and data averages from Kohn, 2010Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
).This study evaluates pCO2-dependencies in the context of two endmember models – a null hypothesis of no dependence vs. a hyperbolic dependence that integrates numerous other studies (Schubert and Jahren, 2012
Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
). Ultimately four datasets are considered: modern leaves (1970 through 2007; Kohn, 2010Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
), tropical rainforest tree rings (least susceptible to changes in MAP; Fig. 2a; van der Sleen et al., 2015van der Sleen, P., Groenendijk, P., Vlam, M., Anten, N.P.R., Boom, A., Bongers, F., Pons, T.L., Terburg, G., Zuidema, P.A. (2015) No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience 8, 24-28.
), sediment organic matter (SOM) from three representative studies of the Pleistocene-Holocene transition (Fig. 2b,c; Hatté et al., 1998Hatté, C., Fontugne, M., Rousseau, D.D., Antoine, P., Zoller, L., Tisnerat-Laborde, N., Bentaleb, I. (1998) δ13C variations of loess organic matter as a record of the vegetation response to climatic changes during the Weichselian. Geology 26, 583-586.
; Sinninghe Damsté et al., 2011Sinninghe Damsté, J.S., Verschuren, D., Ossebaar, J., Blokker, J., van Houten, R., van der Meer, M.T.J., Plessen, B., Schouten, S. (2011) A 25,000-year record of climate-induced changes in lowland vegetation of eastern equatorial Africa revealed by the stable carbon-isotopic composition of fossil plant leaf waxes. Earth and Planetary Science Letters 302, 236-246, doi: 10.1016/j.epsl.2010.12.025.
; Barker et al., 2013Barker, P.A., Hurrell, E.R., Leng, M.J., Plessen, B., Wolff, C., Conley, D.J., Keppens, E., Milne, I., Cumming, B.F., Laird, K.R., Kendrick, C.P., Wynn, P.M., Verschuren, D. (2013) Carbon cycling within an East African lake revealed by the carbon isotope composition of diatom silica: a 25-ka record from Lake Challa, Mt. Kilimanjaro. Quaternary Science Reviews 66, 55-63, doi: 10.1016/j.quascirev.2012.07.016.
), and herbivore collagen and tooth enamel for especially low-pCO2 (<250 ppmv) and high-pCO2 (>500 ppmv) periods of the geologic past (Fig. 3; Supplementary Information). The overall intent of these comparisons is to correct first for factors that we know influence δ13C (especially MAP, carbon sources) and ascribe any residual effect to pCO2.Figure 2 Tree ring and Pleistocene-Holocene records. (a) Tropical data from Thailand show near-zero pCO2 effect. Models are anchored on oldest 5-10 analyses (where any pCO2 effects should be smallest). SJ12 = model of Schubert and Jahren (2012)
Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
. (b) Correction for changes in mean annual precipitation (“ΔMAP”), and possibly for organic matter preservation (“preserv.”) reproduces records independent of a direct pCO2-dependence. Brief climatic events also correlate with changes in Δ13C independent of pCO2. H1, H3 = Heinrich events 1 and 3; DO2 = Dansgaard-Oeschger event 2; B-A = Bølling-Allerød; YD = Younger Dryas. (c) An abrupt shift at c. 15.8 ka could reflect an abrupt threshold response of C4 biomass in the local ecosystem or variable contributions of diatoms to SOM (Fig. S-1).Because plants track the δ13C of atmospheric CO2, which has changed through time, data are presented in terms of isotope discrimination (Δ13C):
Eq. 1
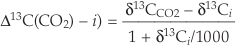
By correcting for secular changes to atmospheric δ13C from recent direct measurements, ice core measurements, and foraminiferal data (Supplementary Information), Equation 1 allows comparisons of data from different times.
Supplementary Information explains materials and methods, including error propagation.
top
Results and Discussion
Modern/Historical records. Linear regression of modern leaf Δ13C (Kohn, 2010
Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
), treating pCO2 as an independent variable (Table S-1), results in a pCO2 coefficient for Δ13C of ‑1.2 ± 1.0 ‰/100 ppmv (2σ). These data do not generally support a positive pCO2-dependence for Δ13C but are quite imprecise. Historical cellulose data from tropical rainforests (Fig. 2a; van der Sleen et al., 2015van der Sleen, P., Groenendijk, P., Vlam, M., Anten, N.P.R., Boom, A., Bongers, F., Pons, T.L., Terburg, G., Zuidema, P.A. (2015) No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience 8, 24-28.
; Table S-1) suggest a positive slope with respect to pCO2 (0.62 ± 1.05 ‰/100 ppmv, 2 s.e.; Supplementary Information), but are also strongly influenced by an up-tick in Δ13C between 2000 and 2010 AD (Fig. 2a; Toona, Chukrasia), which might reflect other factors. For example, 10-20 yr, 1-2 ‰ oscillations at all sites (e.g., Melia, Afzelia, Fig. 2a) probably reflect susceptibility to environmental factors besides precipitation and pCO2, and such an effect might explain specific slopes. Tree ring isotope data across Europe, corrected for climate variables, suggest a negative pCO2-dependence (-0.55 ± 0.67 ‰/100 ppmv; data from Frank et al., 2015Frank, D.C., Poulter, B., Saurer, M., Esper, J., Huntingford, C., Helle, G., Treydte, K., Zimmermann, N.E., Schleser, G.H., Ahlström, A., Ciais, P., Friedlingstein, P., Levis, S., Lomas, M., Sitch, S., Viovy, N., Andreu-Hayles, L., Bednarz, Z., Berninger, F., Boettger, T., D‘Alessandro, C.M., Daux, V., Filot, M., Grabner, M., Gutierrez, E., Haupt, M., Hilasvuori, E., Jungner, H., Kalela-Brundin, M., Krapiec, M., Leuenberger, M., Loader, N.J., Marah, H., Masson-Delmotte, V., Pazdur, A., Pawelczyk, S., Pierre, M., Planells, O., Pukiene, R., Reynolds-Henne, C.E., Rinne, K.T., Saracino, A., Sonninen, E., Stievenard, M., Switsur, V.R., Szczepanek, M., Szychowska-Krapiec, E., Todaro, L., Waterhouse, J.S., Weig, M. (2015) Water-use efficiency and transpiration across European forests during the Anthropocene. Nature Climate Change 5, 579–583, doi: 10.1038/NCLIMATE2614.
). Overall, modern/historical data are too imprecise and idiosyncratic to either require or preclude a pCO2-effect.Pleistocene-Holocene sediment records. Mid-latitude sediments from the Rhine River valley in France and Germany present one of the better cases for a pCO2-effect: a gradual 2 ‰ increase in Δ13C between 20 and 10 ka parallels pCO2 (Fig. 2b). A near doubling of MAP across the Pleistocene-Holocene transition (Bartlein et al., 2011
Bartlein, P.J., Harrison, S.P., Brewer, S., Connor, S., Davis, B.A.S., Gajewski, K., Guiot, J., Harrison-Prentice, T.I., Henderson, A., Peyron, O., Prentice, I.C., Scholze, M., Seppa, H., Shuman, B., Sugita, S., Thompson, R.S., Viau, A.E., Williams, J., Wu, H. (2011) Pollen-based continental climate reconstructions at 6 and 21 ka: a global synthesis. Climate Dynamics 37, 775-802, doi: 10.1007/s00382-010-0904-1.
; Alder and Hostetler, 2015Alder, J.R., Hostetler, S.W. (2015) Global climate simulations at 3000-year intervals for the last 21000 years with the GENMOM coupled atmosphere-ocean model. Climate of the Past 11, 449-471, doi: 10.5194/cp-11-449-2015.
), however, predicts c. 1.25 ‰ of the observed ~2 ‰ increase (Fig. 2b), so any pCO2-effect must be ≤~1.0 ‰/100 ppmv. In addition, organic C content in these records increases by a factor of 9 across the Pleistocene-Holocene boundary (Hatté et al., 1998Hatté, C., Fontugne, M., Rousseau, D.D., Antoine, P., Zoller, L., Tisnerat-Laborde, N., Bentaleb, I. (1998) δ13C variations of loess organic matter as a record of the vegetation response to climatic changes during the Weichselian. Geology 26, 583-586.
). Numerous studies show a linear correlation between Δ13C and the logarithm of soil organic C content, (e.g., Poage and Feng, 2004Poage, M.A., Feng, X. (2004) A theoretical analysis of steady state δ13C profiles of soil organic matter. Global Biogeochemical Cycles, 18, doi: 10.1029/2003GB002195.
) with a minimum slope of c. 0.7 ‰ per log10[C] (Balesdent et al., 1993Balesdent, J., Girardin, C., Mariotti, A. (1993) Site-related δ13C of tree leaves and soil organic matter in a termperate forest. Ecology 74, 1713-1721, doi: 10.2307/1939930.
). If the striking increase in organic C content in the European records represents differential degradation, Δ13C should increase by at least c. 0.65 ‰. The combined isotopic effects of changes to MAP and organic C content (c. 2 ‰) can explain the data without resort to any pCO2-effects.Alternatively, an estimate of ecosystem impacts on Δ13C can be made for the French record in reference to the Dansgaard-Oeschger 2 (DO2) and the Bølling-Allerød (B-A) warming events, which caused brief 2-2.5 ‰ spikes in Δ13C without obvious changes to pCO2. Subtracting this (pCO2-independent) effect from the Pleistocene-Holocene transition yields a residual pCO2-effect of ~0 ‰/100 ppmv.
A large, abrupt increase in Δ13C in low-latitude Kenyan lake data (Fig. 2c) likely reflects decreasing abundances of C4 vegetation (Sinninghe Damsté et al., 2011
Sinninghe Damsté, J.S., Verschuren, D., Ossebaar, J., Blokker, J., van Houten, R., van der Meer, M.T.J., Plessen, B., Schouten, S. (2011) A 25,000-year record of climate-induced changes in lowland vegetation of eastern equatorial Africa revealed by the stable carbon-isotopic composition of fossil plant leaf waxes. Earth and Planetary Science Letters 302, 236-246, doi: 10.1016/j.epsl.2010.12.025.
), rather than increasing pCO2. Increasing pCO2 tends to destabilise C4 plants (Ehleringer et al., 1997Ehleringer, J.R., Cerling, T.E., Helliker, B.R. (1997) C-4 photosynthesis, atmospheric CO2 and climate. Oecologia 112, 285-299, doi: 10.1007/s004420050311.
), so an abrupt decrease in C4 abundance (Prentice et al., 2011Prentice, I.C., Harrison, S.P., Bartlein, P.J. (2011) Global vegetation and terrestrial carbon cycle changes after the last ice age. New Phytologist 189, 988-998.
) during the Pleistocene-Holocene rise in pCO2 logically explains the observations. Changes to lake diatom abundance and preservation may also play a role (Fig. S-1).Overall, for the Pleistocene-Holocene transition, widespread increases in MAP across much of Earth (Bartlein et al., 2011
Bartlein, P.J., Harrison, S.P., Brewer, S., Connor, S., Davis, B.A.S., Gajewski, K., Guiot, J., Harrison-Prentice, T.I., Henderson, A., Peyron, O., Prentice, I.C., Scholze, M., Seppa, H., Shuman, B., Sugita, S., Thompson, R.S., Viau, A.E., Williams, J., Wu, H. (2011) Pollen-based continental climate reconstructions at 6 and 21 ka: a global synthesis. Climate Dynamics 37, 775-802, doi: 10.1007/s00382-010-0904-1.
) and concomitant changes to floral ecosystems (CLIMAP, 1976CLIMAP (1976) The surface of the Ice-Age Earth. Science 191, 1131-1137.
; Prentice et al., 2011Prentice, I.C., Harrison, S.P., Bartlein, P.J. (2011) Global vegetation and terrestrial carbon cycle changes after the last ice age. New Phytologist 189, 988-998.
), especially changes in C3/C4 abundances at low latitudes, generally predict an increase in Δ13C, compromising any quantitative retrieval or validation of pCO2-dependencies using these records. At present, the best constrained data imply a maximum value of ≤1.0 ‰/100 ppmv (MAP-correction alone) or possibly ~0.0 ‰/100 ppmv (DO-event analog, France; MAP plus organic C degradation) between 180 and 270 ppmv.Fossil herbivore records. Pleistocene and Tertiary herbivore data place the most stringent constraints on a pCO2-dependence: ‑0.03 ± 0.13 ‰/100 ppmv between 180 and 370 ppmv, and ‑0.03 ± 0.24 ‰/100 ppmv between 370 and 715 ppmv. The pCO2-dependent model strongly overestimates Δ13C for the Pleistocene and underestimates Δ13C for the Tertiary (Fig. 3a,b). Similarly, isotopically-based estimates of MAP without correction of pCO2 reproduce independent estimates within uncertainties (Fig. 3c), whereas estimates from the pCO2-dependent model strongly overestimate MAP for Pleistocene data, and strongly underestimate MAP for Tertiary data (Fig. 3d). Uncertainties in estimates of MAP, pCO2, or the δ13C of atmospheric CO2 do not likely bias interpretation of Pleistocene data because pCO2 and δ13C are directly determined from ice cores, and MAP is based on large numbers of floral observations coupled to GCM’s with especially well constrained boundary conditions. Large errors for Tertiary pCO2 cause the pCO2-dependent model errors to overlap independent estimates of Δ13C and MAP. The point of overlap, however, occurs only at quasi-modern pCO2 levels, which is generally inconsistent with Eocene and middle Miocene proxies (Beerling and Royer, 2011
Beerling, D.J., Royer, D.L. (2011) Convergent Cenozoic CO2 history. Nature Geoscience 4, 418-420.
). Note that a systematic error for Pleistocene calculations operates in the opposite direction for Tertiary data, so the internal consistency of both sets of calculations (the same pCO2 coefficient) suggests that systematic errors are small.Figure 3 Cenozoic herbivore data. (a-d) Calculated vs. measured Δ13C, and calculated vs. independently estimated MAP, showing good correspondence with pCO2-independent model. Representative error bars (2σ) reflect propagated uncertainties in isotope compositions and tissue-diet fractionations, the dependence of δ13C on MAP, and proxy estimates of MAP and pCO2 (Table S-2).
Considering calculated MAP throughout the Cenozoic, a U-shaped distribution (Fig. 4a) highlights unusually dry conditions [low Δ13C(CO2-plant), high δ13C(plant)] during the late Eocene and early/middle Oligocene. Aggregating Cenozoic MAP estimates, the pCO2-independent model exhibits a predominance of values between 0 and 1.0 m/yr (>80 %; Fig. 4b) with a distribution that mimics modern Earth’s area distribution of MAP (dashed line, Fig. 4b). In contrast, the pCO2-dependent model implies widespread rainforests during the Pleistocene (Fig. 4a), which contrasts starkly with global climate and vegetation syntheses that have long indicated greater aridity across much of the globe (e.g., CLIMAP, 1976
CLIMAP (1976) The surface of the Ice-Age Earth. Science 191, 1131-1137.
; Prentice et al., 2011Prentice, I.C., Harrison, S.P., Bartlein, P.J. (2011) Global vegetation and terrestrial carbon cycle changes after the last ice age. New Phytologist 189, 988-998.
), especially in Europe where many of the Pleistocene fossils were collected. The pCO2-dependent model also implies negative MAP during the Eocene and Oligocene (Fig. 4a). A histogram of pCO2-dependent predictions (Fig. 4c) yields unrealistically abundant hyperarid deserts (MAP < 0) and rainforests (>2.5 m/yr).Figure 4 (a) Estimates of MAP for the Cenozoic based on fossil herbivore isotope compositions show a U-shaped distribution. pCO2-corrections imply widespread rainforests in the Pleistocene, and hyper-deserts in mid-Cenozoic. pCO2 curve simplified from Beerling and Royer, 2011
Beerling, D.J., Royer, D.L. (2011) Convergent Cenozoic CO2 history. Nature Geoscience 4, 418-420.
. (b-c) Histograms of Cenozoic MAP estimates show a better match between the global distribution (by area) of MAP (Kohn, 2010Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
) vs. MAP estimated using the pCO2-independent model. The pCO2-dependent model predicts unusually high proportions of deserts (MAP < 0.1 m/yr) and rainforests (MAP > 2.5 m/yr) in the geologic record.top
Perspective and Recommendations
As discussed elsewhere (Schubert and Jahren, 2012
Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
), studies investigating pCO2-effects rarely account for other factors that may influence δ13C, making cross-study comparisons difficult. One recent study that did attempt to account for such factors (Frank et al., 2015Frank, D.C., Poulter, B., Saurer, M., Esper, J., Huntingford, C., Helle, G., Treydte, K., Zimmermann, N.E., Schleser, G.H., Ahlström, A., Ciais, P., Friedlingstein, P., Levis, S., Lomas, M., Sitch, S., Viovy, N., Andreu-Hayles, L., Bednarz, Z., Berninger, F., Boettger, T., D‘Alessandro, C.M., Daux, V., Filot, M., Grabner, M., Gutierrez, E., Haupt, M., Hilasvuori, E., Jungner, H., Kalela-Brundin, M., Krapiec, M., Leuenberger, M., Loader, N.J., Marah, H., Masson-Delmotte, V., Pazdur, A., Pawelczyk, S., Pierre, M., Planells, O., Pukiene, R., Reynolds-Henne, C.E., Rinne, K.T., Saracino, A., Sonninen, E., Stievenard, M., Switsur, V.R., Szczepanek, M., Szychowska-Krapiec, E., Todaro, L., Waterhouse, J.S., Weig, M. (2015) Water-use efficiency and transpiration across European forests during the Anthropocene. Nature Climate Change 5, 579–583, doi: 10.1038/NCLIMATE2614.
) concluded that pCO2 has no effect on tree δ13C (-0.55 ± 0.67 ‰/100 ppmv; Supplementary Information), in good agreement with results here. Although susceptible to individual idiosyncrasies, cellulose records from individual trees from the Borneo rainforest (Loader et al., 2011Loader, N.J., Walsh, R.P.D., Robertson, I., Bidin, K., Ong, R.C., Reynolds, G., McCarroll, D., Gagen, M., Young, G.H.F. (2011) Recent trends in the intrinsic water-use efficiency of ringless rainforest trees in Borneo. Philosophical Transactions of the Royal Society B-Biological Science 366, 3330-3339.
; MAP c. 3 m/yr) also imply a pCO2-effect indistinguishable from zero (0.0 ± 0.2 ‰/100 ppmv; Supplementary Information). Early Eocene isotopes of angiosperms further support pCO2-independence (Diefendorf et al., 2015Diefendorf, A.F., Freeman, K.H., Wing, S.L., Currano, E.D., Mueller, K.E. (2015) Paleogene plants fractionated carbon isotopes similar to modern plants. Earth and Planetary Science Letters 429, 33-44.
). At present, the most discrepant study (Schubert and Jahren, 2012Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
) represents short-term experiments on two forbs (Arabidopsis and radish). As discussed recently (Diefendorf et al., 2015Diefendorf, A.F., Freeman, K.H., Wing, S.L., Currano, E.D., Mueller, K.E. (2015) Paleogene plants fractionated carbon isotopes similar to modern plants. Earth and Planetary Science Letters 429, 33-44.
; Voelker et al., 2015Voelker, S.L., Brooks, J.R., Meinzer, F.C., Anderson, R., Bader, M.K, Battipaglia, G., Becklin, K.M., Beerling, D., Bert, D., Betancourt, J.L., Dawson, T.E., Domec, J.C., Guyette, R.P., Körner, C., Leavitt, S.W., Linder, S., Marshall, J.D., Mildner, M., Ogée, J., Panyushkina, I., Plumpton, H.J., Pregitzer, K.S., Saurer, M., Smith, A.R., Siegwolf, R.T., Stambaugh, M.C., Talhelm, A.F., Tardif, J.C., van de Water, P.K., Ward, J.K., Wingate, L. (2015) A dynamic leaf gas-exchange strategy is conserved in woody plants under changing ambient CO2: evidence from carbon isotope discrimination in paleo and CO2 enrichment studies. Global Change Biology, doi: 10.1111/gcb13102.
), plants may exhibit short-term phenotypic and isotopic changes in response to abrupt changes to pCO2, but on evolutionary timescales (decades to centuries) may evolve towards an optimal physiology whose isotopic fractionation is pCO2-independent. Short-term experiments might have relevance for modern rapid changes to pCO2, but not for ancient isotope records.Overall, natural data either do not resolve a pCO2-dependence or indicate an effect far smaller than recent models. On geologic timescales, a best estimate for a pCO2-dependence between c. 200 and c. 700 ppmv averages -0.04 ± 0.26 ‰/100 ppmv (2 s.e.; Table S-3), i.e. pCO2-dependencies are negligible. Because no correction for pCO2 is needed, this result vastly simplifies interpretations of past climates and carbon use by C3 land plants.
top
Acknowledgements
Funded by NSF grants EAR1251443 and EAR1349749. Comments from A. Diefendorf and detailed reviews from B. Schubert and an anonymous reviewer helped improve the MS, although Schubert does not necessarily agree with data selection, interpretations, or conclusions.
Editor: Bruce Watson
top
References
Alder, J.R., Hostetler, S.W. (2015) Global climate simulations at 3000-year intervals for the last 21000 years with the GENMOM coupled atmosphere-ocean model. Climate of the Past 11, 449-471, doi: 10.5194/cp-11-449-2015.
 Show in context
Show in context A near doubling of MAP across the Pleistocene-Holocene transition (Bartlein et al., 2011; Alder and Hostetler, 2015), however, predicts c. 1.25 ‰ of the observed ~2 ‰ increase (Fig. 2b), so any pCO2-effect must be ≤~1.0 ‰/100 ppmv.
View in article
Arens, N.C., Jahren, A.H., Amundson, R.G. (2000) Can C3 plants faithfully record the carbon isotopic composition of atmospheric carbon dioxide? Paleobiology 26, 137-164.
Show in context Many studies have argued that increasing pCO2 increases carbon isotope discrimination (e.g., Feng and Epstein, 1995; Schubert and Jahren, 2012); others have argued for no pCO2-effect (e.g., Arens et al., 2000; Saurer et al., 2004; Frank et al., 2015).
View in article
Balesdent, J., Girardin, C., Mariotti, A. (1993) Site-related δ13C of tree leaves and soil organic matter in a termperate forest. Ecology 74, 1713-1721, doi: 10.2307/1939930.
Show in context Numerous studies show a linear correlation between Δ13C and the logarithm of soil organic C content, (e.g., Poage and Feng, 2004) with a minimum slope of c. 0.7 ‰ per log10[C] (Balesdent et al., 1993).
View in article
Barker, P.A., Hurrell, E.R., Leng, M.J., Plessen, B., Wolff, C., Conley, D.J., Keppens, E., Milne, I., Cumming, B.F., Laird, K.R., Kendrick, C.P., Wynn, P.M., Verschuren, D. (2013) Carbon cycling within an East African lake revealed by the carbon isotope composition of diatom silica: a 25-ka record from Lake Challa, Mt. Kilimanjaro. Quaternary Science Reviews 66, 55-63, doi: 10.1016/j.quascirev.2012.07.016.
Show in context Ultimately four datasets are considered: modern leaves (1970 through 2007; Kohn, 2010), tropical rainforest tree rings (least susceptible to changes in MAP; Fig. 2a; van der Sleen et al., 2015), sediment organic matter (SOM) from three representative studies of the Pleistocene-Holocene transition (Fig. 2b,c; Hatté et al., 1998; Sinninghe Damsté et al., 2011; Barker et al., 2013), and herbivore collagen and tooth enamel for especially low-pCO2 (<250 ppmv) and high-pCO2 (>500 ppmv) periods of the geologic past (Fig. 3; Supplementary Information).
View in article
Bartlein, P.J., Harrison, S.P., Brewer, S., Connor, S., Davis, B.A.S., Gajewski, K., Guiot, J., Harrison-Prentice, T.I., Henderson, A., Peyron, O., Prentice, I.C., Scholze, M., Seppa, H., Shuman, B., Sugita, S., Thompson, R.S., Viau, A.E., Williams, J., Wu, H. (2011) Pollen-based continental climate reconstructions at 6 and 21 ka: a global synthesis. Climate Dynamics 37, 775-802, doi: 10.1007/s00382-010-0904-1.
Show in context A near doubling of MAP across the Pleistocene-Holocene transition (Bartlein et al., 2011; Alder and Hostetler, 2015), however, predicts c. 1.25 ‰ of the observed ~2 ‰ increase (Fig. 2b), so any pCO2-effect must be ≤~1.0 ‰/100 ppmv.
View in article
Overall, for the Pleistocene-Holocene transition, widespread increases in MAP across much of Earth (Bartlein et al., 2011) and concomitant changes to floral ecosystems (CLIMAP, 1976; Prentice et al., 2011), especially changes in C3/C4 abundances at low latitudes, generally predict an increase in Δ13C, compromising any quantitative retrieval or validation of pCO2-dependencies using these records.
View in article
Beerling, D.J., Royer, D.L. (2011) Convergent Cenozoic CO2 history. Nature Geoscience 4, 418-420.
Show in context The point of overlap, however, occurs only at quasi-modern pCO2 levels, which is generally inconsistent with Eocene and middle Miocene proxies (Beerling and Royer, 2011).
View in article
Figure 4 [...] pCO2 curve simplified from Beerling and Royer (2011).
View in article
Cao, L., Bala, G., Caldeira, K., Nemani, R., Ban-Weiss, G. (2010) Importance of carbon dioxide physiological forcing to future climate change. Proceedings of the National Academy of Sciences 107, 9513-9518, doi:10.1073/pnas.0913000107.
Show in context C3 plants (trees, shrubs, herbs and cool-climate grasses) constitute c. 95 % of terrestrial plant biomass (Still et al., 2003), so understanding C3 plant response to changes in the partial pressure of atmospheric CO2 (pCO2) is key for modelling Earth’s carbon cycle (e.g., Cao et al., 2010; Shevliakova et al., 2013).
View in article
CLIMAP (1976) The surface of the Ice-Age Earth. Science 191, 1131-1137.
Show in context Overall, for the Pleistocene-Holocene transition, widespread increases in MAP across much of Earth (Bartlein et al., 2011) and concomitant changes to floral ecosystems (CLIMAP, 1976; Prentice et al., 2011), especially changes in C3/C4 abundances at low latitudes, generally predict an increase in Δ13C, compromising any quantitative retrieval or validation of pCO2-dependencies using these records.
View in article
In contrast, the pCO2-dependent model implies widespread rainforests during the Pleistocene (Fig. 4a), which contrasts starkly with global climate and vegetation syntheses that have long indicated greater aridity across much of the globe (e.g., CLIMAP, 1976; Prentice et al., 2011), especially in Europe where many of the Pleistocene fossils were collected.
View in article
Diefendorf, A.F., Mueller, K.E., Wing, S.L., Koch, P.L., Freeman, K.H. (2010) Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proceedings of the National Academy of Sciences 107, 5738-5743.
Show in context Multiple factors besides atmospheric CO2 can affect C3 plant δ13C, including MAP, ontogeny, light levels, genetics and fertilisation (e.g., through atmospheric nitrogen deposition; Farquhar et al., 1989; Stewart et al., 1995; McCarroll and Loader, 2004; Diefendorf et al., 2010; Kohn, 2010; Fig. 1b).
View in article
Other physical factors, including latitude and altitude, have a minor impact on C3 plant δ13C (Diefendorf et al., 2010; Kohn, 2010), but high variability both within and among trees must reflect individual responses to other, less easily identified, environmental or physiological factors (McCarroll and Loader, 2004).
View in article
Diefendorf, A.F., Freeman, K.H., Wing, S.L., Currano, E.D., Mueller, K.E. (2015) Paleogene plants fractionated carbon isotopes similar to modern plants. Earth and Planetary Science Letters 429, 33-44.
Show in context Early Eocene isotopes of angiosperms further support pCO2-independence (Diefendorf et al., 2015).
View in article
As discussed recently (Diefendorf et al., 2015; Voelker et al., 2015), plants may exhibit short-term phenotypic and isotopic changes in response to abrupt changes to pCO2, but on evolutionary timescales (decades to centuries) may evolve towards an optimal physiology whose isotopic fractionation is pCO2-independent.
View in article
Ehleringer, J.R., Cerling, T.E., Helliker, B.R. (1997) C-4 photosynthesis, atmospheric CO2 and climate. Oecologia 112, 285-299, doi: 10.1007/s004420050311.
Show in context Increasing pCO2 tends to destabilise C4 plants (Ehleringer et al., 1997), so an abrupt decrease in C4 abundance (Prentice et al., 2011) during the Pleistocene-Holocene rise in pCO2 logically explains the observations.
View in article
Farquhar, G.D., Ehleringer, J.R., Hubick, K.T. (1989) Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 40, 503-537.
Show in context Carbon isotope compositions (δ13C) of C3 plants are a principal means of inferring water use efficiency (Farquhar et al., 1989), which is important to models of future C3 biomass, and have been proposed to allow estimation of mean annual precipitation (MAP) in the geologic past (Kohn, 2010).
View in article
Multiple factors besides atmospheric CO2 can affect C3 plant δ13C, including MAP, ontogeny, light levels, genetics and fertilisation (e.g., through atmospheric nitrogen deposition; Farquhar et al., 1989; Stewart et al., 1995; McCarroll and Loader, 2004; Diefendorf et al., 2010; Kohn, 2010; Fig. 1b).
View in article
Feng, X., Epstein, S. (1995) Carbon isotopes of trees from arid environments and implications for reconstructing atmospheric CO2 concentration. Geochimica et Cosmochimica Acta 59, 2599-2608.
Show in context Many studies have argued that increasing pCO2 increases carbon isotope discrimination (e.g., Feng and Epstein, 1995; Schubert and Jahren, 2012); others have argued for no pCO2-effect (e.g., Arens et al., 2000; Saurer et al., 2004; Frank et al., 2015).
View in article
Frank, D.C., Poulter, B., Saurer, M., Esper, J., Huntingford, C., Helle, G., Treydte, K., Zimmermann, N.E., Schleser, G.H., Ahlström, A., Ciais, P., Friedlingstein, P., Levis, S., Lomas, M., Sitch, S., Viovy, N., Andreu-Hayles, L., Bednarz, Z., Berninger, F., Boettger, T., D‘Alessandro, C.M., Daux, V., Filot, M., Grabner, M., Gutierrez, E., Haupt, M., Hilasvuori, E., Jungner, H., Kalela-Brundin, M., Krapiec, M., Leuenberger, M., Loader, N.J., Marah, H., Masson-Delmotte, V., Pazdur, A., Pawelczyk, S., Pierre, M., Planells, O., Pukiene, R., Reynolds-Henne, C.E., Rinne, K.T., Saracino, A., Sonninen, E., Stievenard, M., Switsur, V.R., Szczepanek, M., Szychowska-Krapiec, E., Todaro, L., Waterhouse, J.S., Weig, M. (2015) Water-use efficiency and transpiration across European forests during the Anthropocene. Nature Climate Change 5, 579–583, doi: 10.1038/NCLIMATE2614.
Show in context Many studies have argued that increasing pCO2 increases carbon isotope discrimination (e.g., Feng and Epstein, 1995; Schubert and Jahren, 2012); others have argued for no pCO2-effect (e.g., Arens et al., 2000; Saurer et al., 2004; Frank et al., 2015).
View in article
Tree ring isotope data across Europe, corrected for climate variables, suggest a negative pCO2-dependence (-0.55 ± 0.67 ‰/100 ppmv; data from Frank et al., 2015).
View in article
One recent study that did attempt to account for such factors (Frank et al., 2015) concluded that pCO2 has no effect on tree δ13C (-0.55 ± 0.67 ‰/100 ppmv; Supplementary Information), in good agreement with results here.
View in article
Freyer, H.D., Wiesberg, L. (1973) 13C decrease in modern wood due to large-scale combustion of fossil fuels. Naturwissenschaften 60, 517-518.
Show in context Plant δ13C depends on the δ13C of atmospheric CO2 (Freyer and Wiesberg, 1973), but an unresolved decades-long debate centres on whether terrestrial C3 plant δ13C responds to pCO2.
View in article
Hatté, C., Fontugne, M., Rousseau, D.D., Antoine, P., Zoller, L., Tisnerat-Laborde, N., Bentaleb, I. (1998) δ13C variations of loess organic matter as a record of the vegetation response to climatic changes during the Weichselian. Geology 26, 583-586.
Show in context Ultimately four datasets are considered: modern leaves (1970 through 2007; Kohn, 2010), tropical rainforest tree rings (least susceptible to changes in MAP; Fig. 2a; van der Sleen et al., 2015), sediment organic matter (SOM) from three representative studies of the Pleistocene-Holocene transition (Fig. 2b,c; Hatté et al., 1998; Sinninghe Damsté et al., 2011; Barker et al., 2013), and herbivore collagen and tooth enamel for especially low-pCO2 (<250 ppmv) and high-pCO2 (>500 ppmv) periods of the geologic past (Fig. 3; Supplementary Information).
View in article
In addition, organic C content in these records increases by a factor of 9 across the Pleistocene-Holocene boundary (Hatté et al., 1998).
View in article
Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
Show in context Carbon isotope compositions (δ13C) of C3 plants are a principal means of inferring water use efficiency (Farquhar et al., 1989), which is important to models of future C3 biomass, and have been proposed to allow estimation of mean annual precipitation (MAP) in the geologic past (Kohn, 2010).
View in article
Multiple factors besides atmospheric CO2 can affect C3 plant δ13C, including MAP, ontogeny, light levels, genetics and fertilisation (e.g., through atmospheric nitrogen deposition; Farquhar et al., 1989; Stewart et al., 1995; McCarroll and Loader, 2004; Diefendorf et al., 2010; Kohn, 2010; Fig. 1b).
View in article
Other physical factors, including latitude and altitude, have a minor impact on C3 plant δ13C (Diefendorf et al., 2010; Kohn, 2010), but high variability both within and among trees must reflect individual responses to other, less easily identified, environmental or physiological factors (McCarroll and Loader, 2004).
View in article
Figure 1 [...] (b) Mean annual precipitation (data and data averages from Kohn, 2010).
View in article
Ultimately four datasets are considered: modern leaves (1970 through 2007; Kohn, 2010), tropical rainforest tree rings (least susceptible to changes in MAP; Fig. 2a; van der Sleen et al., 2015), sediment organic matter (SOM) from three representative studies of the Pleistocene-Holocene transition (Fig. 2b,c; Hatté et al., 1998; Sinninghe Damsté et al., 2011; Barker et al., 2013), and herbivore collagen and tooth enamel for especially low-pCO2 (<250 ppmv) and high-pCO2 (>500 ppmv) periods of the geologic past (Fig. 3; Supplementary Information).
View in article
Linear regression of modern leaf Δ13C (Kohn, 2010), treating pCO2 as an independent variable (Table S-3), results in a pCO2 coefficient for Δ13C of ‑1.2 ± 1.0 ‰/100 ppmv (2σ).
View in article
Figure 4 [...] (b-c) Histograms of Cenozoic MAP estimates show a better match between the global distribution (by area) of MAP (Kohn, 2010) vs. MAP estimated using the pCO2-independent model.
View in article
Loader, N.J., Walsh, R.P.D., Robertson, I., Bidin, K., Ong, R.C., Reynolds, G., McCarroll, D., Gagen, M., Young, G.H.F. (2011) Recent trends in the intrinsic water-use efficiency of ringless rainforest trees in Borneo. Philosophical Transactions of the Royal Society B-Biological Science 366, 3330-3339.
Show in contextAlthough susceptible to individual idiosyncrasies, cellulose records from individual trees from the Borneo rainforest (Loader et al., 2011; MAP c. 3 m/yr) also imply a pCO2-effect indistinguishable from zero (0.0 ± 0.2 ‰/100 ppmv; Supplementary Information).
View in article
McCarroll, D., Loader, N.J. (2004) Stable isotopes in tree rings. Quaternary Science Reviews 23, 771-801.
Show in context Multiple factors besides atmospheric CO2 can affect C3 plant δ13C, including MAP, ontogeny, light levels, genetics and fertilisation (e.g., through atmospheric nitrogen deposition; Farquhar et al., 1989; Stewart et al., 1995; McCarroll and Loader, 2004; Diefendorf et al., 2010; Kohn, 2010; Fig. 1b).
View in article
Other physical factors, including latitude and altitude, have a minor impact on C3 plant δ13C (Diefendorf et al., 2010; Kohn, 2010), but high variability both within and among trees must reflect individual responses to other, less easily identified, environmental or physiological factors (McCarroll and Loader, 2004).
View in article
Poage, M.A., Feng, X. (2004) A theoretical analysis of steady state δ13C profiles of soil organic matter. Global Biogeochemical Cycles, 18, doi: 10.1029/2003GB002195.
Show in context Numerous studies show a linear correlation between Δ13C and the logarithm of soil organic C content, (e.g., Poage and Feng, 2004) with a minimum slope of c. 0.7 ‰ per log10[C] (Balesdent et al., 1993).
View in article
Prentice, I.C., Harrison, S.P., Bartlein, P.J. (2011) Global vegetation and terrestrial carbon cycle changes after the last ice age. New Phytologist 189, 988-998.
Show in context Increasing pCO2 tends to destabilise C4 plants (Ehleringer et al., 1997), so an abrupt decrease in C4 abundance (Prentice et al., 2011) during the Pleistocene-Holocene rise in pCO2 logically explains the observations.
View in article
Overall, for the Pleistocene-Holocene transition, widespread increases in MAP across much of Earth (Bartlein et al., 2011) and concomitant changes to floral ecosystems (CLIMAP, 1976; Prentice et al., 2011), especially changes in C3/C4 abundances at low latitudes, generally predict an increase in Δ13C, compromising any quantitative retrieval or validation of pCO2-dependencies using these records.
View in article
In contrast, the pCO2-dependent model implies widespread rainforests during the Pleistocene (Fig. 4a), which contrasts starkly with global climate and vegetation syntheses that have long indicated greater aridity across much of the globe (e.g., CLIMAP, 1976; Prentice et al., 2011), especially in Europe where many of the Pleistocene fossils were collected.
View in article
Saurer, M., Siegwolf, R.T.W., Schweingruber, F.H. (2004) Carbon isotope discrimination indicates improving water-use efficiency of trees in northern Eurasia over the last 100 years. Global Change Biology 10, 2109-2120.
Show in context Many studies have argued that increasing pCO2 increases carbon isotope discrimination (e.g., Feng and Epstein, 1995; Schubert and Jahren, 2012); others have argued for no pCO2-effect (e.g., Arens et al., 2000; Saurer et al., 2004; Frank et al., 2015).
View in article
Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
Show in context Many studies have argued that increasing pCO2 increases carbon isotope discrimination (e.g., Feng and Epstein, 1995; Schubert and Jahren, 2012); others have argued for no pCO2-effect (e.g., Arens et al., 2000; Saurer et al., 2004; Frank et al., 2015).
View in article
Recent work (Schubert and Jahren, 2012) integrated natural and experimental data to infer that C3 plant δ13C depends hyperbolically on pCO2 (Fig. 1a) and that changes to carbon isotope compositions in leaves and sediment organic matter over the last 30 ka parallel and predict pCO2 (Schubert and Jahren, 2015).
View in article
Figure 1 [...] Experiments are for above-ground biomass (Schubert and Jahren, 2012), shifted to fit preferred curve.
View in article
This study evaluates pCO2-dependencies in the context of two endmember models – a null hypothesis of no dependence vs. a hyperbolic dependence that integrates numerous other studies (Schubert and Jahren, 2012).
View in article
Figure 2 [...] SJ12 = model of Schubert and Jahren (2012).
View in article
As discussed elsewhere (Schubert and Jahren, 2012), studies investigating pCO2-effects rarely account for other factors that may influence δ13C, making cross-study comparisons difficult.
View in article
At present, the most discrepant study (Schubert and Jahren, 2012) represents short-term experiments on two forbs (Arabidopsis and radish).
View in article
Schubert, B.A., Jahren, A.H. (2015) Global increase in plant carbon isotope fractionation following the Last Glacial Maximum caused by increase in atmospheric pCO2. Geology 43, 435-438.
Show in context Recent work (Schubert and Jahren, 2012) integrated natural and experimental data to infer that C3 plant δ13C depends hyperbolically on pCO2 (Fig. 1a) and that changes to carbon isotope compositions in leaves and sediment organic matter over the last 30 ka parallel and predict pCO2 (Schubert and Jahren, 2015).
View in article
Shevliakova, E., Stouffer, R.J., Malyshev, S., Krasting, J.P., Hurtt, G.C., Pacala, S.W. (2013) Historical warming reduced due to enhanced land carbon uptake. Proceedings of the National Academy of Sciences 110, 16730-16735, doi: 10.1073/pnas.1314047110.
Show in context C3 plants (trees, shrubs, herbs and cool-climate grasses) constitute c. 95 % of terrestrial plant biomass (Still et al., 2003), so understanding C3 plant response to changes in the partial pressure of atmospheric CO2 (pCO2) is key for modelling Earth’s carbon cycle (e.g., Cao et al., 2010; Shevliakova et al., 2013).
View in article
Sinninghe Damsté, J.S., Verschuren, D., Ossebaar, J., Blokker, J., van Houten, R., van der Meer, M.T.J., Plessen, B., Schouten, S. (2011) A 25,000-year record of climate-induced changes in lowland vegetation of eastern equatorial Africa revealed by the stable carbon-isotopic composition of fossil plant leaf waxes. Earth and Planetary Science Letters 302, 236-246, doi: 10.1016/j.epsl.2010.12.025.
Show in context Ultimately four datasets are considered: modern leaves (1970 through 2007; Kohn, 2010), tropical rainforest tree rings (least susceptible to changes in MAP; Fig. 2a; van der Sleen et al., 2015), sediment organic matter (SOM) from three representative studies of the Pleistocene-Holocene transition (Fig. 2b,c; Hatté et al., 1998; Sinninghe Damsté et al., 2011; Barker et al., 2013), and herbivore collagen and tooth enamel for especially low-pCO2 (<250 ppmv) and high-pCO2 (>500 ppmv) periods of the geologic past (Fig. 3; Supplementary Information).
View in article
Increasing pCO2 tends to destabilise C4 plants (Ehleringer et al., 1997), so an abrupt decrease in C4 abundance (Prentice et al., 2011) during the Pleistocene-Holocene rise in pCO2 logically explains the observations.
View in article
Stewart, G.R., Turnbull, M.H., Schmidt, S., Erskine, P.D. (1995) 13C natural abundance in plant communities along a rainfall gradient: a biological integrator of water availability. Australian Journal of Plant Physiology 22, 51-55.
Show in context Multiple factors besides atmospheric CO2 can affect C3 plant δ13C, including MAP, ontogeny, light levels, genetics and fertilisation (e.g., through atmospheric nitrogen deposition; Farquhar et al., 1989; Stewart et al., 1995; McCarroll and Loader, 2004; Diefendorf et al., 2010; Kohn, 2010; Fig. 1b).
View in article
Still, C.J., Berry, J.A., Collatz, G.J., DeFries, R.S. (2003) Global distributions of C3 and C4 vegetation: carbon cycle implications. Global Biogeochemical Cycles 17, doi: 10.1029/2001GB001807.
Show in context C3 plants (trees, shrubs, herbs and cool-climate grasses) constitute c. 95 % of terrestrial plant biomass (Still et al., 2003), so understanding C3 plant response to changes in the partial pressure of atmospheric CO2 (pCO2) is key for modelling Earth’s carbon cycle (e.g., Cao et al., 2010; Shevliakova et al., 2013).
View in article
van der Sleen, P., Groenendijk, P., Vlam, M., Anten, N.P.R., Boom, A., Bongers, F., Pons, T.L., Terburg, G., Zuidema, P.A. (2015) No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience 8, 24-28.
Show in context Ultimately four datasets are considered: modern leaves (1970 through 2007; Kohn, 2010), tropical rainforest tree rings (least susceptible to changes in MAP; Fig. 2a; van der Sleen et al., 2015), sediment organic matter (SOM) from three representative studies of the Pleistocene-Holocene transition (Fig. 2b,c; Hatté et al., 1998; Sinninghe Damsté et al., 2011; Barker et al., 2013), and herbivore collagen and tooth enamel for especially low-pCO2 (<250 ppmv) and high-pCO2 (>500 ppmv) periods of the geologic past (Fig. 3; Supplementary Information).
View in article
Historical cellulose data from tropical rainforests (Fig. 2a; van der Sleen et al., 2015; Table S-3) suggest a positive slope with respect to pCO2 (0.62 ± 1,05 ‰/100 ppmv, 2 s.e.; Supplementary Information), but are also strongly influenced by an up-tick in Δ13C between 2000 and 2010 AD (Fig. 2a; Toona, Chukrasia), which might reflect other factors.
View in article
Voelker, S.L., Brooks, J.R., Meinzer, F.C., Anderson, R., Bader, M.K, Battipaglia, G., Becklin, K.M., Beerling, D., Bert, D., Betancourt, J.L., Dawson, T.E., Domec, J.C., Guyette, R.P., Körner, C., Leavitt, S.W., Linder, S., Marshall, J.D., Mildner, M., Ogée, J., Panyushkina, I., Plumpton, H.J., Pregitzer, K.S., Saurer, M., Smith, A.R., Siegwolf, R.T., Stambaugh, M.C., Talhelm, A.F., Tardif, J.C., van de Water, P.K., Ward, J.K., Wingate, L. (2015) A dynamic leaf gas-exchange strategy is conserved in woody plants under changing ambient CO2: evidence from carbon isotope discrimination in paleo and CO2 enrichment studies. Global Change Biology, doi: 10.1111/gcb13102.
Show in context As discussed recently (Diefendorf et al., 2015; Voelker et al., 2015), plants may exhibit short-term phenotypic and isotopic changes in response to abrupt changes to pCO2, but on evolutionary timescales (decades to centuries) may evolve towards an optimal physiology whose isotopic fractionation is pCO2-independent.
View in article
top
Supplementary Information
SI-1: Methods
SI-1.1 Modern leaf data. For modern leaves, the dataset of Kohn (2010)
Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
was used, adding values for modern changes to pCO2 as determined from recent compilations (www.esrl.noaa.gov/gmd/ccgg/trends/; http://cdiac.esd.ornl.gov/trends/trends.htm; McCarroll and Loader, 2004McCarroll, D., Loader, N.J. (2004) Stable isotopes in tree rings. Quaternary Science Reviews 23, 771-801.
) as an independent variable. These compilations also summarise atmospheric δ13C. Data were regressed vs. latitude, elevation, MAP, and pCO2 (Table S-1). These data span a ~50 ppmv change in pCO2 and integrate thousands of observations, so represent the best leaf dataset for identifying any resolvable pCO2–dependence on timescales of several decades.SI-1.2 Historical tree ring records. For tropical trees, data from van der Sleen et al. (2015)
van der Sleen, P., Groenendijk, P., Vlam, M., Anten, N.P.R., Boom, A., Bongers, F., Pons, T.L., Terburg, G., Zuidema, P.A. (2015) No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience 8, 24-28.
for 27 cm-diameter trees (canopy) were emphasised rather than 8 cm-diameter tree data (understory), because isotopes in closed canopy forest understory can depend on factors such as light intensity, humidity gradients, accumulation of ground-level CO2, and recycling of soil carbon (e.g., Vogel, 1978Vogel, J.C. (1978) Recycling of carbon in a forest environment. Oecologia Plantarum 13, 89-94.
; Medina and Minchin, 1980Medina, E., Minchin, P. (1980) Stratification of δ13C values of leaves in Amazonian rain forests. Oecologia 45, 377-378.
; Ometto et al., 2006Ometto, J.P.H.B., Ehleringer, J.R., Domingues, T.F., Berry, J.A., I., F.Y., Mazzi, E., Higuchi, N., Flanagan, L.B., Nardoto, G.B., Martinelli, L.A. (2006) The stable carbon and nitrogen isotopic composition of vegetation in tropic forests of the Amazon Basin, Brazil. Biogeochemistry 79, 251-274.
). The original dataset includes pCO2 and atmospheric δ13C. This dataset represents a nearly ideal modern/historical test case because it (a) integrates observations in 5-year intervals from scores of each tree species (least susceptible to individual idiosyncrasies), (b) reports exclusively for old-growth forests (least affected by human activities), (c) accounts for ontogeny (all data represent a constant tree diameter), and (d) represents Earth’s wettest environments (least susceptible to changes in MAP: Fig. 1b). If these data do not resolve a pCO2‑dependence or ‑independence, it is difficult to conceive how a better historical dataset could be developed. Isotopic data were ordered according to age and values for Δ13C(CO2-plant) were calculated using records of atmospheric δ13CCO2 as tabulated in the supplemental files of van der Sleen et al. (2015)van der Sleen, P., Groenendijk, P., Vlam, M., Anten, N.P.R., Boom, A., Bongers, F., Pons, T.L., Terburg, G., Zuidema, P.A. (2015) No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience 8, 24-28.
. An unweighted linear least squares method was used to calculate pCO2-dependencies. Canopy data are presented in Figure 2, but understory data do not yield significantly different conclusions.Figure 2a was constructed by comparing values for Δ13C(CO2-plant) to a pCO2-independent model (constant Δ), the hyperbolic pCO2-dependent model (c. 1.5 ‰/100 ppmv between 280 and 390 ppmv; Schubert and Jahren, 2012
Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
), and, for reference, a constant intermediate pCO2-dependence (0.75 ‰/100 ppmv). Because trees could exhibit different species-specific carbon isotope fractionations, the pCO2-dependent models were anchored on Δ13C values for the oldest 5-10 analyses of each record, where pCO2 changes most slowly. Regression results of Δ13C vs. pCO2 for each species and locality were combined into a weighted average and weighted standard error (Table S-1).Table S-1 Specific regression results for leaf and tree ring δ13C.
| Leaf regressions | |||
| Parameter | Value ± 1σ | t | p |
| Intercept | 6.35 ± 2.07 | 3.06 | 0.002 |
| Elevation (m) | -1.80 ± 0.39x10-4 | -4.66 | <1x10-4 |
| log(MAP-300. mm) | 5.84 ± 0.24 | 24.85 | <1x10-4 |
| Abs(latitude. °) | 0.0139 ± 0.0042 | 3.28 | 0.001 |
| pCO2 (ppmv) | -0.0116 ± 0.0052 | -2.23 | 0.026 |
Δ13C = 6.35 - 1.80e-4 x Elev + 5.84*log(MAP-300) + 0.014 x Abs(lat.) – 0.012*pCO2
| Tree ring regressions* | Slope (±2σ) | pCO2 range (ppmv) |
| Thailand A. xylocarpa | 0.10 ± 0.68 | 285-390 |
| Thailand C. tabularis | 0.10 ± 0.54 | 285-385 |
| Thailand M. azedarach | -0.56 ± 1.07 | 300-390 |
| Thailand T. ciliata | 0.80 ± 1.11 | 300-390 |
| Cameroon B. cynometroides | 0.95 ± 0.69 | 300-385 |
| Cameroon B. eurycoma | 0.30 ± 0.83 | 295-375 |
| Cameroon D. ogea | 0.89 ± 0.67 | 280-390 |
| Cameroon T. ivorensis | 1.30 ± 0.53 | 285-385 |
| Bolivia C. ianeirensis | 1.25 ± 0.71 | 300-385 |
| Bolivia H. crepitans | 0.46 ± 1.16 | 290-385 |
| Bolivia S. fruticosa | 0.32 ± 0.70 | 290-390 |
| Average | 0.62 ± 1.05 | 285-390 |
| Borneo S. johorensis† | -0.04 ± 0.20 | 285-385 |
| Borneo S. superba† | 0.14 ± 0.29 | 285-370 |
| European composite† | -0.55 ± 0.67 | 300-370 |
*Regressions for specific tree ring data from van der Sleen et al. (2015)van der Sleen, P., Groenendijk, P., Vlam, M., Anten, N.P.R., Boom, A., Bongers, F., Pons, T.L., Terburg, G., Zuidema, P.A. (2015) No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience 8, 24-28. (Thailand, Cameroon, Bolivia; multiple trees) and Loader et al. (2011Loader, N.J., Walsh, R.P.D., Robertson, I., Bidin, K., Ong, R.C., Reynolds, G., McCarroll, D., Gagen, M., Young, G.H.F. (2011) Recent trends in the intrinsic water-use efficiency of ringless rainforest trees in Borneo. Philosophical Transactions of the Royal Society B-Biological Science 366, 3330-3339.; Borneo; single trees). European composite is from Frank et al. (2015)Frank, D.C., Poulter, B., Saurer, M., Esper, J., Huntingford, C., Helle, G., Treydte, K., Zimmermann, N.E., Schleser, G.H., Ahlström, A., Ciais, P., Friedlingstein, P., Levis, S., Lomas, M., Sitch, S., Viovy, N., Andreu-Hayles, L., Bednarz, Z., Berninger, F., Boettger, T., D‘Alessandro, C.M., Daux, V., Filot, M., Grabner, M., Gutierrez, E., Haupt, M., Hilasvuori, E., Jungner, H., Kalela-Brundin, M., Krapiec, M., Leuenberger, M., Loader, N.J., Marah, H., Masson-Delmotte, V., Pazdur, A., Pawelczyk, S., Pierre, M., Planells, O., Pukiene, R., Reynolds-Henne, C.E., Rinne, K.T., Saracino, A., Sonninen, E., Stievenard, M., Switsur, V.R., Szczepanek, M., Szychowska-Krapiec, E., Todaro, L., Waterhouse, J.S., Weig, M. (2015) Water-use efficiency and transpiration across European forests during the Anthropocene. Nature Climate Change 5, 579–583, doi: 10.1038/NCLIMATE2614. for Quercus and Pinus.
†Data not used in averages.
Results for individual tropical trees (Loader et al., 2011
Loader, N.J., Walsh, R.P.D., Robertson, I., Bidin, K., Ong, R.C., Reynolds, G., McCarroll, D., Gagen, M., Young, G.H.F. (2011) Recent trends in the intrinsic water-use efficiency of ringless rainforest trees in Borneo. Philosophical Transactions of the Royal Society B-Biological Science 366, 3330-3339.
) and for European trees (Frank et al., 2015Frank, D.C., Poulter, B., Saurer, M., Esper, J., Huntingford, C., Helle, G., Treydte, K., Zimmermann, N.E., Schleser, G.H., Ahlström, A., Ciais, P., Friedlingstein, P., Levis, S., Lomas, M., Sitch, S., Viovy, N., Andreu-Hayles, L., Bednarz, Z., Berninger, F., Boettger, T., D‘Alessandro, C.M., Daux, V., Filot, M., Grabner, M., Gutierrez, E., Haupt, M., Hilasvuori, E., Jungner, H., Kalela-Brundin, M., Krapiec, M., Leuenberger, M., Loader, N.J., Marah, H., Masson-Delmotte, V., Pazdur, A., Pawelczyk, S., Pierre, M., Planells, O., Pukiene, R., Reynolds-Henne, C.E., Rinne, K.T., Saracino, A., Sonninen, E., Stievenard, M., Switsur, V.R., Szczepanek, M., Szychowska-Krapiec, E., Todaro, L., Waterhouse, J.S., Weig, M. (2015) Water-use efficiency and transpiration across European forests during the Anthropocene. Nature Climate Change 5, 579–583, doi: 10.1038/NCLIMATE2614.
; their Fig. 2a) are also tabulated for reference, but were not used in calculating weighted mean values because of uncertainties in correcting for climate change (European dataset) and in accounting for ontogeny (individual tropical trees). Data from Loader et al. (2011)Loader, N.J., Walsh, R.P.D., Robertson, I., Bidin, K., Ong, R.C., Reynolds, G., McCarroll, D., Gagen, M., Young, G.H.F. (2011) Recent trends in the intrinsic water-use efficiency of ringless rainforest trees in Borneo. Philosophical Transactions of the Royal Society B-Biological Science 366, 3330-3339.
were regressed similarly to van der Sleen et al. (2015)van der Sleen, P., Groenendijk, P., Vlam, M., Anten, N.P.R., Boom, A., Bongers, F., Pons, T.L., Terburg, G., Zuidema, P.A. (2015) No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience 8, 24-28.
– Δ13C values were calculated from tabulated data and regressed vs. pCO2. The slope for European trees was based on Figure 2a of Frank et al. (2015)Frank, D.C., Poulter, B., Saurer, M., Esper, J., Huntingford, C., Helle, G., Treydte, K., Zimmermann, N.E., Schleser, G.H., Ahlström, A., Ciais, P., Friedlingstein, P., Levis, S., Lomas, M., Sitch, S., Viovy, N., Andreu-Hayles, L., Bednarz, Z., Berninger, F., Boettger, T., D‘Alessandro, C.M., Daux, V., Filot, M., Grabner, M., Gutierrez, E., Haupt, M., Hilasvuori, E., Jungner, H., Kalela-Brundin, M., Krapiec, M., Leuenberger, M., Loader, N.J., Marah, H., Masson-Delmotte, V., Pazdur, A., Pawelczyk, S., Pierre, M., Planells, O., Pukiene, R., Reynolds-Henne, C.E., Rinne, K.T., Saracino, A., Sonninen, E., Stievenard, M., Switsur, V.R., Szczepanek, M., Szychowska-Krapiec, E., Todaro, L., Waterhouse, J.S., Weig, M. (2015) Water-use efficiency and transpiration across European forests during the Anthropocene. Nature Climate Change 5, 579–583, doi: 10.1038/NCLIMATE2614.
, specifically determining their best estimate of ci for years 1900 and 2000 AD, calculating a 2 standard error (s.e.) in ci from the spread of data, ratioing ci to known ca, and calculating Δ13C from standard expressions (Farquhar et al., 1989Farquhar, G.D., Ehleringer, J.R., Hubick, K.T. (1989) Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 40, 503-537.
).SI-1.3 Pleistocene-Holocene sediment records. Numerous records exist for sediment organic matter (SOM) spanning the Pleistocene-Holocene transition, but for simplicity only three localities were considered in detail: loessic sediments from the Rhine River valley in France and Germany (Fig. 2b; Hatté et al., 1998
Hatté, C., Fontugne, M., Rousseau, D.D., Antoine, P., Zoller, L., Tisnerat-Laborde, N., Bentaleb, I. (1998) δ13C variations of loess organic matter as a record of the vegetation response to climatic changes during the Weichselian. Geology 26, 583-586.
) and sediment cores from Lake Challa, Kenya (Fig. 2c; Sinninghe Damsté et al., 2011Sinninghe Damsté, J.S., Verschuren, D., Ossebaar, J., Blokker, J., van Houten, R., van der Meer, M.T.J., Plessen, B., Schouten, S. (2011) A 25,000-year record of climate-induced changes in lowland vegetation of eastern equatorial Africa revealed by the stable carbon-isotopic composition of fossil plant leaf waxes. Earth and Planetary Science Letters 302, 236-246.
; Barker et al., 2013Barker, P.A., Hurrell, E.R., Leng, M.J., Plessen, B., Wolff, C., Conley, D.J., Keppens, E., Milne, I., Cumming, B.F., Laird, K.R., Kendrick, C.P., Wynn, P.M., Verschuren, D. (2013) Carbon cycling within an East African lake revealed by the carbon isotope composition of diatom silica: a 25-ka record from Lake Challa, Mt. Kilimanjaro. Quaternary Science Reviews 66, 55-63.
). These records were chosen as characteristic of mid-latitude and low-latitude sites, and exemplify both the increase in Δ13C(CO2-SOM) commonly observed between c. 20 and c. 10 ka (Schubert and Jahren, 2015Schubert, BA., Jahren, A.H. (2015) Global increase in plant carbon isotope fractionation following the Last Glacial Maximum caused by increase in atmospheric pCO2. Geology 43, 435-438.
), and the issues involved in interpreting these isotopic shifts. Data were converted to Δ13C based on composite ice core δ13CCO2 (Section SI-1.6). Numerous other records are illustrated and discussed in Figure S-1 and Section SI-3.SI-1.4 Fossil herbivore records. For fossil herbivore collagen and tooth enamel, a literature search retrieved 57 studies representing 98 discrete time intervals and locations (Table S-2) with sufficient isotopic and chronologic resolution to permit calculations of MAP. This dataset omits studies with a suspected C4 bias, represents a substantial increase over the 22 studies discussed in Kohn (2010)
Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
, and particularly emphasises low-pCO2 (Pleistocene) and high-pCO2 (mid-Miocene and Eocene) times in the context of independently estimated MAP, pCO2, and the δ13C of atmospheric CO2. The compilation is intended to be illustrative rather than exhaustive. Emphasis was placed on studies of multiple species to minimise taxon-specific dietary and isotopic biases, so studies of just one species were avoided to minimise potential species-specific biases. Data for Pleistocene caribou were omitted because consumption of high δ13C lichens biases their compositions relative to C3 consumers (Fox-Dobbs et al., 2008Fox-Dobbs, K., Leonard, J.A., Koch, P.L. (2008) Pleistocene megafauna from eastern Beringia: paleoecological and paleoenvironmental interpretations of stable carbon and nitrogen isotope and radiocarbon records. Palaeogeography, Palaeoclimatology, Palaeoecology 261, 30-46.
). If sources reported a mean composition for all herbivores, then that value was used. Otherwise a median value, rounded to the nearest 0.25 ‰ was estimated. Uncertainties in mean or median values are typically ~±0.5 ‰ (2 s.e.), and that value has been used for error propagation.Table S-2 Herbivore isotope data and calculations of Δ13C and MAP.
| Source | Age (Ma) | δ13C (CO2. ‰) | pCO2 ppmv | δ13C (‰) | δ13C (plant. ‰) | Δ13C (K)* | Δ13C (S)* | Alt. (m) | Lat. (˚) | MAP (mm/yr) | Δ13C (K)* | Δ13C (S)* | MAP (K)* | MAP (S)* |
| Aiglstorfer et al. (2014) Aiglstorfer, M., Bocherens, H., Bohme, M. (2014) Large mammal ecology in the late Middle Miocene Gratkorn locality (Austria). Palaeobiodiversity and Palaeoenvironments 94, 189-213. | 12.1 | -6.1 | 300 | -12 | -26.25 | 20.69 | 21.79 | 350 | 47 | 919 | 1571 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 4 | -6.25 | 350 | -10 | -24.5 | 18.71 | 18.99 | 2000 | 35 | 377 | 456 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 6 | -6.2 | 300 | -9.75 | -23.75 | 17.98 | 19.07 | 2000 | 35 | 208 | 480 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 7 | -6.15 | 300 | -10 | -24 | 18.29 | 19.38 | 2000 | 35 | 274 | 581 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 7.5 | -6.1 | 300 | -9.5 | -23.5 | 17.82 | 18.91 | 2000 | 35 | 178 | 433 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 9 | -5.95 | 300 | -9.5 | -24 | 18.49 | 19.59 | 2000 | 35 | 322 | 655 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 9.5 | -6 | 300 | -10.5 | -24.5 | 18.96 | 20.06 | 2000 | 35 | 448 | 848 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 11.5 | -6 | 300 | -9.5 | -23.5 | 17.92 | 19.02 | 2000 | 35 | 197 | 463 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 13 | -5.85 | 300 | -10 | -24 | 18.6 | 19.69 | 2000 | 35 | 348 | 694 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 14 | -5.75 | 325 | -9.5 | -23.5 | 18.18 | 18.84 | 2000 | 35 | 250 | 414 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 17 | -5.4 | 550 | -9 | -23 | 18.01 | 16.19 | 2000 | 35 | 216 | -47 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 25 | -6 | 400 | -10 | -24 | 18.44 | 18.06 | 2000 | 35 | 310 | 225 | |||
| Biasatti et al. (2012) Biasatti, D., Wang, Y., Gao, F., Xu, Y., Flynn, L. (2012) Paleoecologies and paleoclimates of late Cenozoic mammals from Southwest China: evidence from stable carbon and oxygen isotopes. Journal of Asian Earth Science 44, 48-61. | 4 | -6.25 | 350 | -12 | -26 | 20.28 | 20.56 | 2000 | 25 | 1016 | 1169 | |||
| Biasatti et al. (2012) Biasatti, D., Wang, Y., Gao, F., Xu, Y., Flynn, L. (2012) Paleoecologies and paleoclimates of late Cenozoic mammals from Southwest China: evidence from stable carbon and oxygen isotopes. Journal of Asian Earth Science 44, 48-61. | 5 | -6.3 | 400 | -12 | -26 | 20.23 | 19.84 | 2000 | 25 | 990 | 809 | |||
| Biasatti et al. (2012) Biasatti, D., Wang, Y., Gao, F., Xu, Y., Flynn, L. (2012) Paleoecologies and paleoclimates of late Cenozoic mammals from Southwest China: evidence from stable carbon and oxygen isotopes. Journal of Asian Earth Science 44, 48-61. | 7.5 | -6.1 | 300 | -12.5 | -27 | 21.48 | 22.57 | 2000 | 25 | 1807 | 2934 | |||
| Biasatti et al. (2012) Biasatti, D., Wang, Y., Gao, F., Xu, Y., Flynn, L. (2012) Paleoecologies and paleoclimates of late Cenozoic mammals from Southwest China: evidence from stable carbon and oxygen isotopes. Journal of Asian Earth Science 44, 48-61. | 8 | -5.95 | 300 | -13 | -27 | 21.63 | 22.73 | 2000 | 25 | 1938 | 3136 | |||
| Bibi (2007) Bibi, F. (2007) Dietary niche partitioning among fossil bovids in late Miocene C3 habitats: Consilience of functional morphology and stable isotope analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 253, 529-538. | 8 | -5.95 | 300 | -10.75 | -25.25 | 19.8 | 20.89 | 450 | 33 | 630 | 1127 | |||
| Blondel et al. (1997) Blondel, C., Bocherens, H., Mariotti, A. (1997) Stable carbon and oxygen isotope ratios in ungulate teeth from French Eocene and Oligocene localities. Bulletin Societé Geologique de France 168, 775-781. | 29.5 | -6.3 | 540 | -9.8 | -24.05 | 18.19 | 16.44 | 50 | 44 | 153 | -71 | |||
| Blondel et al. (1997) Blondel, C., Bocherens, H., Mariotti, A. (1997) Stable carbon and oxygen isotope ratios in ungulate teeth from French Eocene and Oligocene localities. Bulletin Societé Geologique de France 168, 775-781. | 33 | -6 | 850 | -9 | -23.25 | 17.66 | 14.25 | 50 | 44 | 69 | -203 | |||
| Blondel et al. (1997) Blondel, C., Bocherens, H., Mariotti, A. (1997) Stable carbon and oxygen isotope ratios in ungulate teeth from French Eocene and Oligocene localities. Bulletin Societé Geologique de France 168, 775-781. | 35 | -6 | 950 | -9.9 | -24.15 | 18.6 | 14.85 | 50 | 44 | 233 | -178 | |||
| Blondel et al. (1997) Blondel, C., Bocherens, H., Mariotti, A. (1997) Stable carbon and oxygen isotope ratios in ungulate teeth from French Eocene and Oligocene localities. Bulletin Societé Geologique de France 168, 775-781. | 37.5 | -6.2 | 880 | -9.3 | -23.55 | 17.77 | 14.25 | 50 | 44 | 85 | -203 | |||
| Boardman and Secord (2013) Boardman, G.S., Secord, R. (2013) Stable isotope paleoecology of White River ungulates during the Eocene-Oligocene climate transition in northwestern Nebraska. Palaeogeography, Palaeoclimatology, Palaeoecology 375, 38-49. | 33 | -5.25 | 850 | -8.9 | -23.15 | 18.32 | 14.91 | 500 | 43 | 198 | -169 | |||
| Boardman and Secord (2013) Boardman, G.S., Secord, R. (2013) Stable isotope paleoecology of White River ungulates during the Eocene-Oligocene climate transition in northwestern Nebraska. Palaeogeography, Palaeoclimatology, Palaeoecology 375, 38-49. | 35 | -6 | 950 | -9.4 | -23.65 | 18.08 | 14.32 | 500 | 43 | 152 | -196 | |||
| Bocherens and Drucker (2003) Bocherens, H., Drucker, D. (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from Recent and ancient terrestrial ecosystems. International Journal of Osteoarcheology 13, 46-53. | 0.02 | -6.6 | 195 ± 10 | -20 | -25 | 18.87 | 22.42 | 50 | 44 | 250 ± 100 | 18.68 | 22.23 | 293 | 2075 |
| Bocherens et al. (2011) Bocherens, H., Drucker, D.G., Bonjean, D., Bridault, A., Conard, N.J., Cupillard, C., Germonpre, M., Honeisen, M., Munzel, S.C., Napierala, H., Patou-Mathis, M., Stephan, E., Uerpmann, H.P., Ziegler, R. (2011) Isotopic evidence for dietary ecology of cave lion (Panthera spelaea) in North-Western Europe: Prey choice, competition and implications for extinction. Quaternary International 245, 249-261. | 0.013 | -6.8 | 240 ± 10 | -20.5 | -25.5 | 19.19 | 21.53 | 1000 | 47 | 550 ± 200 | 19.64 | 21.98 | 412 | 1477 |
| Bocherens et al. (2011) Bocherens, H., Drucker, D.G., Bonjean, D., Bridault, A., Conard, N.J., Cupillard, C., Germonpre, M., Honeisen, M., Munzel, S.C., Napierala, H., Patou-Mathis, M., Stephan, E., Uerpmann, H.P., Ziegler, R. (2011) Isotopic evidence for dietary ecology of cave lion (Panthera spelaea) in North-Western Europe: Prey choice, competition and implications for extinction. Quaternary International 245, 249-261. | 0.013 | -6.8 | 240 ± 10 | -20.75 | -25.75 | 19.45 | 21.79 | 75 | 49 | 400 ± 200 | 19.36 | 21.69 | 426 | 1514 |
| Bocherens et al. (2011) Bocherens, H., Drucker, D.G., Bonjean, D., Bridault, A., Conard, N.J., Cupillard, C., Germonpre, M., Honeisen, M., Munzel, S.C., Napierala, H., Patou-Mathis, M., Stephan, E., Uerpmann, H.P., Ziegler, R. (2011) Isotopic evidence for dietary ecology of cave lion (Panthera spelaea) in North-Western Europe: Prey choice, competition and implications for extinction. Quaternary International 245, 249-261. | 0.034 | -6.7 | 210 ± 10 | -20.5 | -25.5 | 19.29 | 22.4 | 300 | 50 | 300 ± 100 | 18.93 | 22.04 | 391 | 2035 |
| Bocherens et al. (2011) Bocherens, H., Drucker, D.G., Bonjean, D., Bridault, A., Conard, N.J., Cupillard, C., Germonpre, M., Honeisen, M., Munzel, S.C., Napierala, H., Patou-Mathis, M., Stephan, E., Uerpmann, H.P., Ziegler, R. (2011) Isotopic evidence for dietary ecology of cave lion (Panthera spelaea) in North-Western Europe: Prey choice, competition and implications for extinction. Quaternary International 245, 249-261. | 0.034 | -6.7 | 210 ± 10 | -20.25 | -25.25 | 19.03 | 22.14 | 750 | 47 | 550 ± 200 | 19.69 | 22.8 | 356 | 1916 |
| Bocherens et al. (2015a) Bocherens, H., Drucker, D.G., Germonpre, M., Laznickova-Galetova, M., Naito, Y.I., Wissing, C., Bruzek, J., Oliva, M. (2015a) Reconstruction of the Gravettian food-web at Predmosti I using multi-isotopic tracking (C-13, N-15, S-34) of bone collagen. Quaternary International 359, 211-228. | 0.0305 | -6.7 | 205 ± 10 | -20.75 | -25.75 | 19.55 | 22.8 | 250 | 49 | 400 ± 200 | 19.32 | 22.57 | 466 | 2437 |
| Bocherens et al. (2015b) Bocherens, H., Hofman-Kaminska, E., Drucker, D.G., Schmolcke, U., Kowalczyk, R. (2015b) European Bison as a refugee species? Evidence from isotopic data on early Holocene Bison and other large herbivores in northern Europe. PLoS One 10, 10.1371/journal.pone.0115090. | 0.011 | -6.6 | 280 | -21 | -26 | 19.92 | 21.39 | 20 | 55 | 542 | 1199 | |||
| Britton et al. (2012) Britton, K., Gaudzinski-Windheuser, S., Roebroeks, W., Kindler, L., Richards, M.P. (2012) Stable isotope analysis of well-preserved 120,000-year-old herbivore bone collagen from the Middle Palaeolithic site of Neumark-Nord 2, Germany reveals niche separation between bovids and equids. Palaeogeography, Palaeoclimatology, Palaeoecology 333, 168-177. | 0.12 | -6.3 | 270 | -21.5 | -26.5 | 20.75 | 22.42 | 125 | 51 | 901 | 2011 | |||
| Castaños et al. (2014) Castaños, J., Zuluaga, M.C., Ortega, L.A., Murelaga, X., Alonso-Olazabal, A., Rofes, J., Castaños, P. (2014) Carbon and nitrogen stable isotopes of bone collagen of large herbivores from the Late Pleistocene Kiputz IX cave site (Gipuzkoa, north Iberian Peninsula) for palaeoenvironmental reconstruction. Quaternary International 339, 131-138. | 0.021 | -6.6 | 195 ± 10 | -21 | -26 | 19.92 | 23.46 | 200 | 43 | 550 ± 200 | 19.75 | 23.29 | 608 | 3337 |
| Cerling et al. (1997) Cerling, T.E., Harris, J.M., MacFadden, B.J., Leakey, M.G., Quade, J., Eisenmann, V., Ehleringer, J.R. (1997) Global vegetation change through the Miocene/Pliocene boundary. Nature 389, 153-158. | 14 | -5.75 | 325 | -10.5 | -24.5 | 19.22 | 19.89 | 1500 | 0 | 650 | 933 | |||
| Coltrain et al. (2004) Coltrain, J.B., Harris, J.M., Cerling, T.E., Ehleringer, J.R., Dearing, M.-D., Ward, J., Allen, J. (2004) Rancho La Brea stable isotope biogeochemistry and its implications for the palaeoecology of late Pleistocene, coastal Southern California. Palaeogeography, Palaeoclimatology, Palaeoecology 205, 199-219. | 0.012 | -6.7 | 265 | -21 | -26 | 19.82 | 21.59 | 50 | 34 | 602 | 1508 | |||
| Coltrain et al. (2004) Coltrain, J.B., Harris, J.M., Cerling, T.E., Ehleringer, J.R., Dearing, M.-D., Ward, J., Allen, J. (2004) Rancho La Brea stable isotope biogeochemistry and its implications for the palaeoecology of late Pleistocene, coastal Southern California. Palaeogeography, Palaeoclimatology, Palaeoecology 205, 199-219. | 0.0225 | -6.7 | 190 ± 10 | -20.5 | -25.5 | 19.29 | 22.99 | 50 | 34 | 450 ± 200 | 19.34 | 23.04 | 435 | 2827 |
| DeSantis et al. (2008) DeSantis, L.R.G., Wallace, S.C. (2008) Neogene forests from the Appalachians of Tennessee, USA: Geochemical evidence from fossil mammal teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 266, 59-68. | 5.75 | -6.2 | 300 | -13.5 | -27.5 | 21.9 | 23 | 900 | 36 | 1860 | 3015 | |||
| DeSantis et al. (2009) DeSantis, L.R.G., Feranec, R.S., MacFadden, B.J. (2009) Effects of Global Warming on Ancient Mammalian Communities and Their Environments. PLoS One 4, 10.1371/journal.pone.0005750. | 1.8 | -6.6 | 250 | -12.5 | -27 | 20.97 | 23.07 | 50 | 29 | 1152 | 3009 | |||
| Domingo et al. (2012) Domingo, L., Koch, P.L., Grimes, S.T., Morales J., Lopez-Martinez N. (2012) Isotopic paleoecology of mammals and the Middle Miocene Cooling event in the Madrid Basin (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 339, 98-113. | 14 | -5.75 | 325 | -9.75 | -23.75 | 18.44 | 19.11 | 1000 | 40 | 249 | 413 | |||
| Domingo et al. (2012) Domingo, L., Koch, P.L., Grimes, S.T., Morales J., Lopez-Martinez N. (2012) Isotopic paleoecology of mammals and the Middle Miocene Cooling event in the Madrid Basin (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 339, 98-113. | 15.5 | -5.25 | 550 ± 250 | -9 | -23 | 18.17 | 16.35 | 1000 | 40 | 300 ± 100 | 18.66 | 16.84 | 194 | -58 |
| Domingo et al. (2013) Domingo, L., Koch, P.L., Fernandez, M.H., Fox, D.L., Domingo, M.S., Alberdi, M.T. (2013) Late Neogene and early Quaternary paleoenvironmental and paleoclimatic conditions in southwestern Europe: Isotopic analyses on mammalian taxa. PLoS One 8, doi: 10.1371/journal.pone.0063739. | 9.5 | -6 | 300 | -10.5 | -24.75 | 19.23 | 20.32 | 1000 | 40 | 448 | 848 | |||
| Eberle et al. (2009) Eberle, J., Fricke, H., Humphrey, J. (2009) Lower-latitude mammals as year-round residents in Eocene Arctic forests. Geology 37, 499-502. | 52.5 | -5.9 | 715 ± 300 | -12 | -26 | 20.64 | 17.8 | 250 | 77 | 1300 ± 400 | 21.79 | 18.96 | 717 | 35 |
| Ecker et al. (2013) Ecker, M., Bocherens, H., Julien, M.A., Rivals, F., Raynal, J.P., Moncel, M.H. (2013) Middle Pleistocene ecology and Neanderthal subsistence: Insights from stable isotope analyses in Payre (Ardeche, southeastern France). Journal of Human Evolution 65, 363-373. | 0.175 | -7 | 200 | -12.5 | -27 | 20.55 | 23.95 | 200 | 45 | 856 | 4070 | |||
| Feranec and MacFadden (2006) Feranec, R.S., MacFadden, B.J. (2006) Isotopic discrimination of resource partitioning among ungulates in C3-dominated communities from the Miocene of Florida and California. Paleobiology 32, 191-205. | 9.5 | -6 | 300 | -11.7 | -25.7 | 20.22 | 21.31 | 50 | 33 | 762 | 1330 | |||
| Feranec et al. (2010) Feranec, R.S., Garcia, N., Diez, J.C., Arsuaga, J.L. (2010) Understanding the ecology of mammalian carnivorans and herbivores from Valdegoba cave (Burgos, northern Spain) through stable isotope analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 263-272. | 0.04 | -6.7 | 200 | -10.3 | -24.55 | 18.3 | 21.69 | 1000 | 42 | 215 | 1647 | |||
| Fox-Dobbs et al. (2008) Fox-Dobbs, K., Leonard, J.A., Koch, P.L. (2008) Pleistocene megafauna from eastern Beringia: paleoecological and paleoenvironmental interpretations of stable carbon and nitrogen isotope and radiocarbon records. Palaeogeography, Palaeoclimatology, Palaeoecology 261, 30-46. | 0.03 | -6.7 | 200 ± 10 | -21 | -26 | 19.82 | 23.21 | 300 | 65 | 275 ± 100 | 19.02 | 22.41 | 486 | 2672 |
| France et al. (2007) France, C.A.M., Zelanko, P.M., Kaufman, A.J., Holtz, T.R. (2007) Carbon and nitrogen isotopic analysis of Pleistocene mammals from the Saltville Quarry (Virginia, USA): Implications for trophic relationships. Palaeogeography, Palaeoclimatology, Palaeoecology 249, 271-282. | 0.016 | -6.8 | 230 ± 10 | -21 | -26 | 19.71 | 22.29 | 500 | 37 | 650 ± 200 | 19.9 | 22.48 | 584 | 2128 |
| Gilmour (2011) Gilmour, D.M. (2011) Chronology and ecology of late Pleistocene megafauna in the northern Willamette Valley, Oregon. MS (Portland State University, Portland, Oregon). | 0.014 | -6.8 | 240 | -21 | -26 | 19.71 | 22.05 | 50 | 45 | 520 | 1746 | |||
| Iacumin et al. (1997) Iacumin, P., Bocherens, H., Delgado Huertas, A., Mariotti, A., Longinelli, A. (1997) A stable isotope study of fossil mammal remains from the Paglicci Cave, southern Italy: N and C as palaeoenvironmental indicators. Earth and Planetary Science Letters 148, 349-357. | 0.023 | -6.7 | 195 ± 10 | -20.5 | -25.5 | 19.29 | 22.84 | 100 | 41 | 550 ± 200 | 19.74 | 23.29 | 412 | 2553 |
| Koch et al. (1995) Koch, P.L., Zachos, J.C., Dettman, D.L. (1995) Stable isotope stratigraphy and paleoclimatology of the Paleogene Bighorn Basin (Wyoming, USA). Palaeogeography, Palaeoclimatology, Palaeoecology 115, 61-89. | 54 | -5.5 | 600 ± 300 | -12.5 | -26.5 | 21.57 | 19.4 | 500 | 44 | 1300 ± 400 | 21.32 | 19.14 | 1467 | 454 |
| Koch et al. (1995) Koch, P.L., Zachos, J.C., Dettman, D.L. (1995) Stable isotope stratigraphy and paleoclimatology of the Paleogene Bighorn Basin (Wyoming, USA). Palaeogeography, Palaeoclimatology, Palaeoecology 115, 61-89. | 57 | -4.3 | 420 | -10 | -24 | 20.18 | 19.57 | 500 | 44 | 726 | 505 | |||
| Kocsis et al. (2014) Kocsis, L., Ozsvart, P., Becker, D., Ziegler, R., Scherler, L., Codrea, V. (2014) Orogeny forced terrestrial climate variation during the late Eocene-early Oligocene in Europe. Geology 42, 727-730. | 31 | -6 | 650 | -11.5 | -25.75 | 20.27 | 17.79 | 500 | 47 | 746 | 96 | |||
| Kocsis et al. (2014) Kocsis, L., Ozsvart, P., Becker, D., Ziegler, R., Scherler, L., Codrea, V. (2014) Orogeny forced terrestrial climate variation during the late Eocene-early Oligocene in Europe. Geology 42, 727-730. | 36 | -6 | 920 | -8.5 | -22.75 | 17.14 | 13.48 | 500 | 47 | 7 | -227 | |||
| Kohn and McKay (2012) Kohn, M.J., McKay, M.P. (2012) Paleoecology of late Pleistocene-Holocene faunas of eastern and central Wyoming, USA, with implications for LGM climate models. Palaeogeography, Palaeoclimatology, Palaeoecology 326-328, 42-53. | 0.02 | -6.6 | 195 ± 10 | -10 | -24.5 | 18.35 | 21.89 | 1500 | 45 | 200 ± 100 | 18.16 | 21.71 | 238 | 1855 |
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 23.1 | -5.9 | 300 | -11.55 | -25.55 | 20.16 | 21.25 | 100 | 45 | 681 | 1205 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 20.5 | -6.1 | 300 | -11.98 | -25.98 | 20.41 | 21.5 | 100 | 45 | 781 | 1359 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 27.5 | -6.3 | 450 | -10.22 | -24.22 | 18.36 | 17.42 | 100 | 45 | 185 | 36 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 30.3 | -6.1 | 650 | -11.27 | -25.27 | 19.66 | 17.18 | 100 | 45 | 507 | 5 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 33.6 | -5.25 | 875 | -10.54 | -24.54 | 19.78 | 16.27 | 100 | 45 | 543 | -86 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 38.4 | -6.2 | 850 | -11.15 | -25.15 | 19.44 | 16.02 | 100 | 45 | 439 | -106 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 39.5 | -6 | 850 | -11.36 | -25.36 | 19.86 | 16.45 | 100 | 45 | 572 | -71 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 42 | -5.7 | 950 | -10.65 | -24.65 | 19.43 | 15.68 | 100 | 45 | 436 | -131 | |||
| Kuitems et al. (2015) Kuitems, M., van der Plicht, J., Druckerc, D.G., van Kolfschotena, T., Palstrab, S.W.L., Bocherens, H. (2015) Carbon and nitrogen stable isotopes of well-preserved Middle Pleistocene bone collagen from Schöningen (Germany) and their palaeoecological implications. Journal of Human Evolution, doi: 10.1016/j.jhevol.2015.1001.1008. | 0.285 | -7 | 225 | -21.25 | -26.25 | 19.77 | 22.48 | 150 | 52 | 515 | 2054 | |||
| Kuitems et al. (2015) Kuitems, M., van der Plicht, J., Druckerc, D.G., van Kolfschotena, T., Palstrab, S.W.L., Bocherens, H. (2015) Carbon and nitrogen stable isotopes of well-preserved Middle Pleistocene bone collagen from Schöningen (Germany) and their palaeoecological implications. Journal of Human Evolution, doi: 10.1016/j.jhevol.2015.1001.1008. | 0.3 | -6.9 | 275 | -22.25 | -27.25 | 20.92 | 22.49 | 150 | 52 | 979 | 2066 | |||
| Macfadden and Higgins (2004) MacFadden, B.J., Higgins, P. (2004) Ancient ecology of 15-million-year-old browsing mammals within C3 plant communities from Panama. Oecologia 140, 169-182. | 16 | -5.25 | 550 | -13 | -27 | 22.35 | 20.53 | 20 | 9 | 2459 | 1052 | |||
| Martin et al. (2011) Martin, C., Bentaleb, I., Antoine, P.O. (2011) Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 19-29. | 9 | -5.9 | 300 | -11 | -25 | 19.59 | 20.68 | 500 | 29 | 577 | 1046 | |||
| Martin et al. (2011) Martin, C., Bentaleb, I., Antoine, P.O. (2011) Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 19-29. | 13 | -5.9 | 300 | -11 | -25 | 19.59 | 20.68 | 500 | 29 | 577 | 1046 | |||
| Martin et al. (2011) Martin, C., Bentaleb, I., Antoine, P.O. (2011) Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 19-29. | 23 | -5.9 | 300 | -10.5 | -24.5 | 19.07 | 20.16 | 500 | 29 | 415 | 797 | |||
| Martin et al. (2011) Martin, C., Bentaleb, I., Antoine, P.O. (2011) Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 19-29. | 29 | -6.3 | 500 | -10.25 | -24.25 | 18.4 | 16.98 | 500 | 29 | 249 | 16 | |||
| Maung-Thein et al. (2011) Maung-Thein, Z.-M. Takai, M., Uno, H., Wynn, J.G., Egi, N., Tsubamoto, T., Thaung-Htike, Aung-Naing-Soe, Maung-Maung, Nishimura, T., Yoneda, M. (2011) Stable isotope analysis of the tooth enamel of Chaingzauk mammalian fauna (late Neogene, Myanmar) and its implication to paleoenvironment and paleogeography. Palaeogeography, Palaeoclimatology, Palaeoecology 300, 11-22. | 6 | -6.2 | 300 | -11 | -25.5 | 19.81 | 20.9 | 300 | 22 | 676 | 1197 | |||
| Merceron et al. (2006) Merceron, G., Zazzo, A., Spassov, N., Geraads, D., Kovachev, D. (2006) Bovid paleoecology and paleoenvironments from the Late Miocene of Bulgaria: evidence from dental microwear and stable isotopes. Palaeogeography, Palaeoclimatology, Palaeoecology 241, 637-654. | 7.85 | -6 | 300 | -12 | -26.5 | 21.06 | 22.15 | 400 | 41 | 1155 | 1934 | |||
| Montanari et al. (2013) Montanari, S., Louys, J., Price, G.J. (2013) Pliocene paleoenvironments of southeastern Queensland, Australia inferred from stable isotopes of marsupial tooth enamel. PLoS One 8, doi: 10.1371/journal.pone.0066221. | 3.4 | -6.5 | 350 | -12.4 | -24.4 | 18.35 | 18.63 | 350 | 27 | 238 | 301 | |||
| Morgan et al. (1994) Morgan, M.E., Kingston, J.D., Marino, B.D. (1994) Carbon isotopic evidence for the emergence of C4 plants in the Neogene from Pakistan and Kenya. Nature 367, 162-165. | 10 | -6 | 300 | -12.5 | -26.5 | 21.06 | 22.15 | 450 | 34 | 1217 | 2029 | |||
| Nelson (2007) Nelson, S.V. (2007) Isotopic reconstructions of habitat change surrounding the extinction of Sivapithecus, a Miocene hominoid, in the Siwalik Group of Pakistan. Palaeogeography, Palaeoclimatology, Palaeoecology 243, 204-222. | 9 | -5.95 | 300 | -11.25 | -25.25 | 19.8 | 20.89 | 400 | 33 | 626 | 1121 | |||
| Passey et al. (2002) Passey, B.H., Cerling, T.E., Perkins, M.E., Voorhies, M.R., Harris, J.M., Tucker, S.T. (2002) Environmental change in the Great Plains; an isotopic record from fossil horses. Journal of Geology 110, 123-140. | 10 | -6 | 300 | -10 | -24 | 18.44 | 19.54 | 800 | 38 | 247 | 540 | |||
| Pushkina et al. (2014) Pushkina, D., Bocherens, H., Ziegler, R. (2014) Unexpected palaeoecological features of the Middle and Late Pleistocene large herbivores in southwestern Germany revealed by stable isotopic abundances in tooth enamel. Quaternary International 339, 164-178. | 0.035 | -6.7 | 200 ± 10 | -11.75 | -26 | 19.82 | 23.21 | 250 | 50 | 300 ± 100 | 18.93 | 22.33 | 547 | 2902 |
| Pushkina et al. (2014) Pushkina, D., Bocherens, H., Ziegler, R. (2014) Unexpected palaeoecological features of the Middle and Late Pleistocene large herbivores in southwestern Germany revealed by stable isotopic abundances in tooth enamel. Quaternary International 339, 164-178. | 0.6 | -6.7 | 250 | -13.5 | -28 | 21.91 | 24.02 | 250 | 50 | 1627 | 4092 | |||
| Qu et al. (2014) Qu, Y.T., Jin, C.Z., Zhang, Y.Q., Hu, Y.W., Shang, X., Wang, C.S. (2014) Preservation assessments and carbon and oxygen isotopes analysis of tooth enamel of Gigantopithecus blacki and contemporary animals from Sanhe Cave, Chongzuo, South China during the Early Pleistocene. Quaternary International 354, 52-58. | 1.4 | -6.6 | 250 | -15.5 | -28.5 | 22.54 | 24.65 | 200 | 22 | 2521 | 6131 | |||
| Raghavan et al. (2014) Raghavan, M., Themudo, G.E., Smith, C.I., Zazula, G., Campos, P.F. (2014) Musk ox (Ovibos moschatus) of the mammoth steppe: tracing palaeodietary and palaeoenvironmental changes over the last 50,000 years using carbon and nitrogen isotopic analysis. Quaternary Science Reviews 102, 192-201. | 0.021 | -6.6 | 195 ± 10 | -20.25 | -25.25 | 19.13 | 22.68 | 100 | 75 | 225 ± 100 | 18.95 | 22.5 | 264 | 1958 |
| Raghavan et al. (2014) Raghavan, M., Themudo, G.E., Smith, C.I., Zazula, G., Campos, P.F. (2014) Musk ox (Ovibos moschatus) of the mammoth steppe: tracing palaeodietary and palaeoenvironmental changes over the last 50,000 years using carbon and nitrogen isotopic analysis. Quaternary Science Reviews 102, 192-201. | 0.055 | -6.7 | 210 | -20.25 | -25.25 | 19.03 | 22.14 | 100 | 75 | 241 | 1529 | |||
| Richards and Hedges (2003) Richards, M.P., Hedges, R.E.M. (2003) Variations in bone collagen δ13C and d15N values of fauna from Northwest Europe over the last 40000 years. Palaeogeography, Palaeoclimatology, Palaeoecology 193, 261-267. | 0.035 | -6.7 | 200 ± 10 | -20 | -25 | 18.77 | 22.16 | 100 | 50 | 350 ± 100 | 19.18 | 22.57 | 254 | 1796 |
| Scherler et al. (2014) Scherler, L., Tütken, T., Becker, D. (2014) Carbon and oxygen stable isotope compositions of late Pleistocene mammal teeth from dolines of Ajoie (Northwestern Switzerland). Quaternary Research 82, 378-387. | 0.035 | -6.7 | 200 ± 10 | -11.6 | -25.6 | 19.4 | 22.79 | 600 | 48 | 450 ± 200 | 19.41 | 22.8 | 446 | 2521 |
| Scherler et al. (2014) Scherler, L., Tütken, T., Becker, D. (2014) Carbon and oxygen stable isotope compositions of late Pleistocene mammal teeth from dolines of Ajoie (Northwestern Switzerland). Quaternary Research 82, 378-387. | 0.08 | -6.5 | 220 | -11.5 | -25.5 | 19.5 | 22.34 | 500 | 48 | 470 | 2041 | |||
| Secord et al. (2008) Secord, R., Wing, S.L., Chew, A. (2008) Stable isotopes in early Eocene mammals as indicators of forest canopy structure and resource partitioning. Paleobiology 34, 282-300. | 53.75 | -5.6 | 620 ± 300 | -13 | -27 | 21.99 | 19.69 | 500 | 44 | 1300 ± 400 | 21.32 | 19.02 | 1784 | 546 |
| Secord et al. (2010) Secord, R., Wing, S.L., Chew, A. (2008) Stable isotopes in early Eocene mammals as indicators of forest canopy structure and resource partitioning. Paleobiology 34, 282-300. | 55.8 | -4.8 | 460 ± 150 | -11 | -25 | 20.72 | 19.68 | 500 | 44 | 1350 ± 400 | 21.4 | 20.35 | 965 | 541 |
| Secord et al. (2010) Secord, R., Wing, S.L., Chew, A. (2008) Stable isotopes in early Eocene mammals as indicators of forest canopy structure and resource partitioning. Paleobiology 34, 282-300. | 57 | -4.3 | 420 | -11 | -25 | 21.23 | 20.61 | 500 | 44 | 1246 | 913 | |||
| Segalen and Lee-Thorp (2009) Ségalen, L., Lee-Thorp, J.A. (2009) Palaeoecology of late Early Miocene fauna in the Namib based on 13C/12C and 18O/16O ratios of tooth enamel and ratite eggshell carbonate. Palaeogeography, Palaeoclimatology, Palaeoecology 277, 191-198. | 17.5 | -5.6 | 400 | -10 | -24 | 18.85 | 18.47 | 100 | 27 | 343 | 253 | |||
| Suraprasit et al. (2014) Suraprasit, K., Chaimanee, Y., Bocherens, H., Chavasseau, O., Jaeger, J.J. (2014) Systematics and phylogeny of middle Miocene Cervidae (Mammalia) from Mae Moh Basin (Thailand) and a paleoenvironmental estimate using enamel isotopy of sympatric herbivore species. Journal of Vertebrate Paleontology 34, 179-194. | 13.3 | -5.7 | 300 | -10.5 | -25 | 19.79 | 20.89 | 500 | 19 | 702 | 1238 | |||
| Tütken and Vennemann (2009) Tütken, T., Vennemann, T. (2009) Stable isotope ecology of Miocene large mammals from Sandelzhausen, southern Germany. Paläontologische Zeitschrift 83, 207-226. | 16.25 | -5.3 | 550 ± 250 | -11.5 | -25.5 | 20.73 | 18.91 | 450 | 49 | 850 ± 250 | 20.55 | 18.73 | 933 | 304 |
| Tütken et al. (2006) Tütken, T., Vennemann, T.W., Janz, H., Heizmann, E.P.J. (2006) Palaeoenvironment and palaeoclimate of the Middle Miocene lake in the Steinheim basin, SW Germany: a reconstruction from C, O, and Sr isotopes of fossil remains. Palaeogeography, Palaeoclimatology, Palaeoecology 241, 457-491. | 13.5 | -5.85 | 300 | -11.1 | -25.1 | 19.75 | 20.84 | 600 | 49 | 549 | 1003 | |||
| Tütken et al. (2013) Tütken, T., Kaiser, T.M., Vennemann, T., Merceron, G. (2013) Opportunistic feeding strategy for the earliest Old World hypsodont equids: Evidence from stable isotope and dental wear proxies. PLoS One 8, doi: 10.1371/journal.pone.0074463. | 10 | -6 | 300 ± 50 | -12.6 | -26.6 | 21.16 | 22.26 | 200 | 48 | 1200 ± 400 | 21.27 | 22.36 | 1141 | 1912 |
| Uno et al. (2011) Uno, K.T., Cerling, T.E., Harris, J.M., Kunimatsu, Y., Leakey, M.G., Nakatsukasa, M., Nakaya, H. (2011) Late Miocene to Pliocene carbon isotope record of differential diet change among East African herbivores. Proceedings of the National Academy of Sciences 108, 6509-6514. | 9.9 | -6 | 300 | -9.4 | -23.9 | 18.34 | 19.43 | 1500 | 1 | 369 | 726 | |||
| van Dam and Reichart (2009) van Dam, J.A., Reichart, G.J. (2009) Oxygen and carbon isotope signatures in late Neogene horse teeth from Spain and application as temperature and seasonality proxies. Palaeogeography, Palaeoclimatology, Palaeoecology 274, 64-81. | 3 | -6.4 | 350 | -11.5 | -25.5 | 19.6 | 19.88 | 900 | 40 | 559 | 659 | |||
| van Dam and Reichart (2009) van Dam, J.A., Reichart, G.J. (2009) Oxygen and carbon isotope signatures in late Neogene horse teeth from Spain and application as temperature and seasonality proxies. Palaeogeography, Palaeoclimatology, Palaeoecology 274, 64-81. | 7.5 | -6.1 | 300 | -11 | -25 | 19.38 | 20.48 | 900 | 40 | 490 | 912 | |||
| Wang and Deng (2005) Wang, Y., Deng, T. (2005) A 25 m.y. isotopic record of paleodiet and environmental change from fossil mammals and paleosols from the NE margin of the Tibetan Plateau. Earth and Planetary Science Letters 236, 322-338. | 10 | -6 | 300 | -10 | -24 | 18.44 | 19.54 | 2000 | 35 | 310 | 636 | |||
| White et al. (2009) White, T.D., Ambrose, S.H., Suwa, G., Su, D.F., DeGusta, D., Bernor, R.L., Boisserie, J.-R., Brunet, M., Delson, E., Frost, S., Garcia, N., Giaourtsakis, I.X., Haile-Selassie, Y., Howell, F.C., Lehmann, T., Likius, A., Pehlevan, C., Saegusa, H., Semprebon, G., Teaford, M., Vrba, E. (2009) Macrovertebrate paleontology and the Pliocene habitat of Ardipithecus ramidus. Science 326, 87-93. | 4.4 | -6.2 | 350 | -11.5 | -25.5 | 19.81 | 20.09 | 1600 | 9 | 849 | 984 | |||
| Zanazzi and Kohn (2008) Zanazzi, A., Kohn, M.J. (2008) Ecology and physiology of White River mammals based on stable isotope ratios of teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 257, 22-37. | 33 | -5.25 | 850 | -9.25 | -23.25 | 18.43 | 15.01 | 500 | 43 | 219 | -164 | |||
| Zanazzi and Kohn (2008) Zanazzi, A., Kohn, M.J. (2008) Ecology and physiology of White River mammals based on stable isotope ratios of teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 257, 22-37. | 35 | -6 | 950 | -10 | -24 | 18.44 | 14.69 | 500 | 43 | 221 | -180 | |||
| Zhang et al. (2012) Zhang, C.F., Wang, Y., Li, Q., Wang, X.M., Deng, T., Tseng, Z.J., Takeuchi, G.T., Xie, G.P., Xu, Y.F. (2012) Diets and environments of late Cenozoic mammals in the Qaidam Basin, Tibetan Plateau: Evidence from stable isotopes. Earth and Planetary Science Letters 333, 70-82. | 11.25 | -6 | 300 | -10.5 | -24.5 | 18.96 | 20.06 | 3000 | 37 | 500 | 928 |
*K = model of Kohn (2010)Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695. (pCO2-independent); S = model of Schubert and Jahren (2012)Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43. (pCO2-dependent)
Note: Uncertainties in δ13C of atmospheric CO2 for the pre-Pleistocene is ~±0.35 ‰ (Tipple et al., 2010Tipple, B.J., Meyers, S.R., Pagani, M. (2010) Carbon isotope ratio of Cenozoic CO2: a comparative evaluation of available geochemical proxies. Paleoceanography 25, doi: 10.1029/2009PA001851.).
Plant δ13C values were inferred from either the mean or median isotope compositions of herbivores (typically ±0.5 ‰, 2 s.e.). Herbivore-diet fractionations for tooth enamel assumed a 14 ‰ offset for perissodactyls (Cerling and Harris, 1999
Cerling, T.E., Harris, J.M. (1999) Carbon isotope fractionation between diet and bioapatite in ungulate mammals and implications for ecological and paleoecological studies. Oecologia 120, 347-363.
) and notoungulates (Kohn et al., 2015Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37.
) and 14.5 ‰ for artiodactyls (Passey et al., 2005Passey, B.H., Robinson, T.F., Ayliffe, L.K., Cerling, T.E., Sponheimer, M., Dearing, M.D., Roeder, B.L., Ehleringer, J.R. (2005) Carbon isotope fractionation between diet, breath CO2, and bioapatite in different mammals. Journal of Archaeological Science 32, 1459-1470.
). For mixed faunas (both perissodactyls and artiodactyls), a 14.25 ‰ offset was applied. An uncertainty of ±0.25 ‰ for this correction was assumed. For collagen, a 5 ‰ offset was applied, relevant to wild herbivores consuming C3 plants, with an assumed uncertainty of ±0.5 ‰ (see summary of Froehle et al., 2010Froehle, A.W., Kellner, C.M., Schoeninger, M.J. (2010) FOCUS: effect of diet and protein source on carbon stable isotope ratios in collagen: follow up to Warinner and Tuross (2009). Journal of Archaeological Science 37, 2662-2670.
). Data were converted to Δ13C (Fig. 3a,b) based on δ13CCO2 inferred from ice core and foraminiferal measurements (Section SI-1.6). Calculated Δ13C for the pCO2-independent model was based on equation 2 of Kohn (2010)Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
using MAP, altitude and latitude as input parameters. For the pCO2-dependent model, these Δ13C values were then corrected according to Schubert and Jahren (2012)Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
.Herbivore data represent highly integrated measures of leaf δ13C: if a 50 kg herbivore (e.g., sheep or goat), consumes c. 2 kg of leaves per day (Kohn, 1996
Kohn, M.J. (1996) Predicting animal ä18O: Accounting for diet and physiological adaptation. Geochimica et Cosmochimica Acta 60, 4811-4829.
), and each leaf weighs 0.5 g (c. 0.05 cm x 2 cm x 5 cm), a single bulk tooth enamel isotope composition that integrates 1-2 years of diet (Balasse et al., 2003Balasse, M., Smith, A.B., Ambrose, S.H., Leigh, S.R. (2003) Determining sheep birth seasonality by analysis of tooth enamel oxygen isotopes ratios: the late Stone Age site of Kasteelberg (South Africa). Journal of Archaeological Science 30, 205-215.
) would represent approximately 2 million leaves.SI-1.5 Estimates of MAP. Independent MAP estimates were taken from regional floral studies (Wilf, 2000
Wilf, P. (2000) Late Paleocene-early Eocene climate changes in southwestern Wyoming: paleobotanical analysis. Geological Society of America Bulletin 112, 292-307.
; Coltrain et al., 2004Coltrain, J.B., Harris, J.M., Cerling, T.E., Ehleringer, J.R., Dearing, M.-D., Ward, J., Allen, J. (2004) Rancho La Brea stable isotope biogeochemistry and its implications for the palaeoecology of late Pleistocene, coastal Southern California. Palaeogeography, Palaeoclimatology, Palaeoecology 205, 199-219.
; Greenwood et al., 2010Greenwood, D.R., Basinger, J.F., Smith, R.Y. (2010) How wet was the Arctic Eocene rain forest? Estimates of precipitation from Paleogene Arctic macrofloras. Geology 38, 15-18.
; Bartlein et al., 2011Bartlein, P.J., Harrison, S.P., Brewer, S., Connor, S., Davis, B.A.S., Gajewski, K., Guiot, J., Harrison-Prentice, T.I., Henderson, A., Peyron, O., Prentice, I.C., Scholze, M., Seppa, H., Shuman, B., Sugita, S., Thompson, R.S., Viau, A.E., Williams, J., Wu, H. (2011) Pollen-based continental climate reconstructions at 6 and 21 ka: a global synthesis. Climate Dynamics 37, 775-802.
; Böhme, 2011Böhme, M., Winklhofer, M., Ilg A. (2011) Miocene precipitation in Europe: temporal trends and spatial gradients. Palaeogeography, Palaeoclimatology, Palaeoecology 304, 212-218.
; Melles et al., 2012Melles, M., Brigham-Grette, J., Minyuk, P.S., Nowaczyk, N.R., Wennrich, V., DeConto, R.M., Anderson, P.M., Andreev, A.A., Coletti, A., Cook, T.L., Haltia-Hovi, E., Kukkonen, M., Lozhkin, A.V., Rosen, P., Tarasov, P., Vogel, H., Wagner, B. (2012) 2.8 Million Years of Arctic Climate Change from Lake El'gygytgyn, NE Russia. Science 337, 315-320.
), and/or from general circulation models (Henrot et al., 2010Henrot, A.J., François, L., Favre, E., Butzin, M., Ouberdous, M., Munhoven, G. (2010) Effects of CO2, continental distribution, topography and vegetation changes on the climate of the Middle Miocene: a model study. Climate of the Past 6, 675-694.
; Krapp and Jungclaus, 2011Krapp, M., Jungclaus, J.H. (2011) The Middle Miocene climate as modelled in an atmosphere-ocean-biosphere model. Climate of the Past 7, 1169-1188.
; Alder and Hostetler, 2015Alder, J.R., Hostetler, S.W. (2015) Global climate simulations at 3000-year intervals for the last 21000 years with the GENMOM coupled atmosphere-ocean model. Climate of the Past 11, 449-471.
). Errors in MAP were taken from original sources and range from ±100 to ±400 mm/yr. Isotopically-based MAP (Figs. 3c,d, 4) used the model of Kohn (2010)Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
, either directly (pCO2-independent) or after first correcting compositions for potential pCO2-dependence (Schubert and Jahren, 2012Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
) based on independently measured or estimated pCO2.SI-1.6 Corrections for atmospheric δ13C and pCO2. For modern data (last few centuries), pCO2 and δ13C are commonly provided in the supplemental files of the original publications, and are based on direct measures of atmospheric CO2 and its isotope composition (McCarroll and Loader, 2004
McCarroll, D., Loader, N.J. (2004) Stable isotopes in tree rings. Quaternary Science Reviews 23, 771-801.
; www.esrl.noaa.gov/gmd/ccgg/trends/; http://cdiac.esd.ornl.gov/trends/trends.htm). For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999Francey, R.J., Allison, C.E., Etheridge, D.M., Trudinger, C.M., Enting, I.G., Leuenberger, M., Langenfelds, R.L., Michel, E., Steele, L.P. (1999) A 1000-year high precision record of δ13C in atmospheric CO2. Tellus Series B 51, 170-193.
; Indermühle et al., 1999Indermühle, A., Stocker, T.F., Joos, F., Fischer, H., Smith, H.J., Wahlen, M., Deck, B., Mastroianni, D., Tschumi, J., Blunier, T., Meyer, R., Stauffer, B. (1999) Holocene carbon-cycle dynamics based on CO2 trapped in ice at Taylor Dome, Antarctica. Nature 398, 121-126.
, 2000Indermühle, A., Monnin, E., Stauffer, B., Stocker, T.F., Wahlen, M. (2000) Atmospheric CO2 concentration from 60 to 20 kyr BP from the Taylor Dome ice core, Antarctica. Geophysical Research Letters 27, 735-738.
; Petit et al., 1999Petit, J.R., Jouzel, J., Raynaud, D., Barkov, N.I., Barnola, J.M., Basile, I., Bender, M., Chappellaz, J., Davis, M., Delaygue, G., Delmotte, M., Kotlyakov, V.M., Legrand, M., Lipenkov, V.Y., Lorius, C., Pepin, L., Ritz, C., Saltzman, E., Stievenard, M. (1999) Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica. Nature 399, 429-436.
; Smith et al., 1999Smith, H.J., Fischer, H., Wahlen, M., Mastroianni, D., Deck, B. (1999) Dual modes of the carbon cycle since the Last Glacial Maximum. Nature 400, 248-250.
; Siegenthaler et al., 2005Siegenthaler, U., Stocker, T.F., Monnin, E., Lüthi, D., Schwander, J., Stauffer, B., Raynaud, D., Barnola, J.M., Fischer, H., Masson-Delmotte, V., Jouzel, J. (2005) Stable carbon cycle-climate relationship during the late Pleistocene. Science 310, 1313-1317.
; Lüthi et al., 2008Lüthi, D., Le Floch, M., Bereiter, B., Blunier, T., Barnola, J.M., Siegenthaler, U., Raynaud, D., Jouzel, J., Fischer, H., Kawamura, K., Stocker, T.F. (2008) High-resolution carbon dioxide concentration record 650,000-800,000 years before present. Nature 453, 379-382.
; Elsig et al., 2009Elsig, J., Schmitt, J., Leuenberger, D., Schneider, R., Eyer, M., Leuenberger, M., Joos, F., Fischer, H., Stocker, T.F. (2009) Stable isotope constraints on Holocene carbon cycle changes from an Antarctic ice core. Nature 461, 507-510.
; Lourantou et al., 2010Lourantou, A., Lavric, J.V., Kohler, P., Barnola, J.M., Paillard, D., Michel, E., Raynaud, D., Chappellaz, J. (2010) Constraint of the CO2 rise by new atmospheric carbon isotopic measurements during the last deglaciation. Global Biogeochemical Cycles 24, doi: 10.1029/2009gb003545.
; Bereiter et al., 2012Bereiter, B., Lüthi, D., Siegrist, M., Schupbach, S., Stocker, T.F., Fischer, H. (2012) Mode change of millennial CO2 variability during the last glacial cycle associated with a bipolar marine carbon seesaw. Proceedings of the National Academy of Sciences 109, 9755-9760.
; Pedro et al., 2012Pedro, J.B., Rasmussen, S.O., van Ommen, T.D. (2012) Tightened constraints on the time-lag between Antarctic temperature and CO2 during the last deglaciation. Climate of the Past 8, 1213-1221.
). For data older than 3 Ma, pCO2 and δ13C values were estimated from geochemical proxies (Seki et al., 2010Seki, O., Foster, G.L., Schmidt, D.N., Mackensen, A., Kawamura, K., Pancost, R.D. (2010) Alkenone and boron-based Pliocene pCO2 records. Earth and Planetary Science Letters 292, 201-211.
; Tipple et al., 2010Tipple, B.J., Meyers, S.R., Pagani, M. (2010) Carbon isotope ratio of Cenozoic CO2: a comparative evaluation of available geochemical proxies. Paleoceanography 25, doi: 10.1029/2009PA001851.
; Beerling and Royer, 2011Beerling, D.J., Royer, D.L. (2011) Convergent Cenozoic CO2 history. Nature Geoscience 4, 418-420.
); the 3 Ma filtered estimates for δ13C of Tipple et al. (2010)Tipple, B.J., Meyers, S.R., Pagani, M. (2010) Carbon isotope ratio of Cenozoic CO2: a comparative evaluation of available geochemical proxies. Paleoceanography 25, doi: 10.1029/2009PA001851.
were used, except for rapid changes across the Eocene-Oligocene transition, for which a more precise estimate was based on high-resolution foraminiferal records (Coxall et al., 2005Coxall, H.K., Wilson, P.A., Pälike, H., Lear, C.H., Backman, J. (2005) Rapid stepwise onset of Antarctic glaciation and deeper calcite compensation in the Pacific Ocean. Nature 433, 53-57.
). Errors in pCO2 for the late Pleistocene were assumed to be ±10 ppmv (as large or larger than the reproducibility of measurements from ice cores). Errors in pCO2 for Tertiary records were assigned based on variation in alternate proxies (±150-300 ppmv; Table S-2). Errors in δ13C were assumed to be ≤0.1 ‰ for the Pleistocene, and ±0.35 ‰ for older materials (Tipple et al., 2010Tipple, B.J., Meyers, S.R., Pagani, M. (2010) Carbon isotope ratio of Cenozoic CO2: a comparative evaluation of available geochemical proxies. Paleoceanography 25, doi: 10.1029/2009PA001851.
).SI-1.7 Error propagation. Propagated errors in Δ13C and MAP include the following contributions:
pCO2: ±10 ppmv for Pleistocene; ±150-300 ppmv for Tertiary.
Diet-tissue fractionation in herbivores: ±0.5 ‰ for collagen; ±0.25 ‰ for tooth enamel.
Herbivore tissue mean/median values: ±0.5 ‰.
δ13C atmospheric CO2: <0.1 ‰ for Pleistocene, ±0.35 ‰ for Tertiary
MAP: ±100-400 mm/yr (uncertainties generally decrease with lower MAP).
Errors were propagated individually by perturbing the preferred value of an input parameter by its 2σ uncertainty, calculating a new derived parameter value, and differencing with the original derived parameter value. For example, the effect of pCO2 uncertainties on Δ13C (for the pCO2-dependent model) would be propagated as follows for Pleistocene data: (a) the calculated value for Pleistocene Δ13C would be calculated using the preferred value for pCO2 (say 290 ppmv: “Δ13C290”), (b) pCO2 would be perturbed by 10 ppmv (i.e. 300 ppmv), (c) Δ13C would be recalculated at 300 ppmv (“Δ13C300”), and (d) the two values for Δ would be differenced (Δ13C290 - Δ13C300). Total errors were determined quadratically, i.e. by taking the square root of the sum of the squares of the propagated component errors. All errors are reported at either 2σ (individual regressions) or 2 s.e. (pooled data).
SI-2: Example Calculation – France et al. (2007)
France, C.A.M., Zelanko, P.M., Kaufman, A.J., Holtz, T.R. (2007) Carbon and nitrogen isotopic analysis of Pleistocene mammals from the Saltville Quarry (Virginia, USA): Implications for trophic relationships. Palaeogeography, Palaeoclimatology, Palaeoecology 249, 271-282.
Collagen data for the Saltville site were first averaged for each species, then, after omitting Rangifer (caribou), a median δ13C value of ‑21.0 ‰ was determined. Plant δ13C was calculated by subtracting 5 ‰ [δ13C (plant) = -26.0 ‰]. For the age of the site (16 ka) an atmospheric δ13C value of -6.8 ‰ was estimated from ice core records (Smith et al., 1999
Smith, H.J., Fischer, H., Wahlen, M., Mastroianni, D., Deck, B. (1999) Dual modes of the carbon cycle since the Last Glacial Maximum. Nature 400, 248-250.
; Lourantou et al., 2010Lourantou, A., Lavric, J.V., Kohler, P., Barnola, J.M., Paillard, D., Michel, E., Raynaud, D., Chappellaz, J. (2010) Constraint of the CO2 rise by new atmospheric carbon isotopic measurements during the last deglaciation. Global Biogeochemical Cycles 24, doi: 10.1029/2009gb003545.
), yielding Δ13C (CO2-plant) = 19.7 ‰. For the site’s altitude (c. 500 m) and latitude (37 °N), equation 2 of Kohn (2010)Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
yields an apparent MAP of 584 mm/yr. Application of equation 6 of Schubert and Jahren (2012)Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
, accounting for lower pCO2 (230 ppmv; Pedro et al., 2012Pedro, J.B., Rasmussen, S.O., van Ommen, T.D. (2012) Tightened constraints on the time-lag between Antarctic temperature and CO2 during the last deglaciation. Climate of the Past 8, 1213-1221.
) relative to reference conditions (AD 2000, pCO2 = 370 ppmv) implies corrected Δ13C = 22.3 ‰, or MAP = 2130 mm/yr. The increases in Δ and MAP reflect principles illustrated in Figure 1: the proposed correction for low pCO2 (230 ppmv) relative to reference conditions at AD 2000 (370 ppmv) decreases δ13Cplant, increases Δ13C, and increases calculated MAP.Independent estimates of Pleistocene MAP are commonly presented as a difference relative to modern day precipitation, which was estimated from online sources at ~1150 mm/yr (Johnson City, TN). Floral studies (Bartlein et al., 2011
Bartlein, P.J., Harrison, S.P., Brewer, S., Connor, S., Davis, B.A.S., Gajewski, K., Guiot, J., Harrison-Prentice, T.I., Henderson, A., Peyron, O., Prentice, I.C., Scholze, M., Seppa, H., Shuman, B., Sugita, S., Thompson, R.S., Viau, A.E., Williams, J., Wu, H. (2011) Pollen-based continental climate reconstructions at 6 and 21 ka: a global synthesis. Climate Dynamics 37, 775-802.
) and the PMIP3 composite GCM results (Alder and Hostetler, 2015Alder, J.R., Hostetler, S.W. (2015) Global climate simulations at 3000-year intervals for the last 21000 years with the GENMOM coupled atmosphere-ocean model. Climate of the Past 11, 449-471.
) both imply ΔMAP of ~-500 mm/yr during the Pleistocene, i.e. Pleistocene MAP = 650 mm/yr. Using this value as independent input to equation 2 of Kohn (2010)Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
, an expected Δ13C value of 19.9 ‰ was calculated for this latitude and altitude. Equation 6 of Schubert and Jahren (2012)Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
implies an additional ~2.6 ‰ correction, yielding an expected Δ13C value of 22.5 ‰. The value determined from isotope measurements is 19.7 ‰.SI-3: Pleistocene-Holocene Transition – Further Discussion
Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015)
Schubert, BA., Jahren, A.H. (2015) Global increase in plant carbon isotope fractionation following the Last Glacial Maximum caused by increase in atmospheric pCO2. Geology 43, 435-438.
, but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994Van de Water, P.K., Leavitt, S.W., Betancourt, J.L. (1994) Trends in stomatal density and 13C/12C ratios of Pinus flexilis needles during last Glacial-Interglacial cycle. Science 264, 239-243.
; Beerling, 1996Beerling, D.J. (1996) 13C discrimination by fossil leaves during the late-glacial climate oscillation 12-10 ka BP: Measurements and physiological controls. Oecologia 108, 29-37.
; Huang et al., 1996Huang, Y.S., Bol, R., Harkness, D.D., Ineson, P., Eglinton, G. (1996) Post-glacial variations in distributions, C-13 and C-14 contents of aliphatic hydrocarbons and bulk organic matter in three types of British acid upland soils. Organic Geochemistry 24, 273-287.
; Hatté et al., 1998Hatté, C., Fontugne, M., Rousseau, D.D., Antoine, P., Zoller, L., Tisnerat-Laborde, N., Bentaleb, I. (1998) δ13C variations of loess organic matter as a record of the vegetation response to climatic changes during the Weichselian. Geology 26, 583-586.
; Beerling and Rundgren, 2000Beerling, D.J., Rundgren, M. (2000) Leaf metabolic and morphological responses of dwarf willow (Salix herbacea) in the sub-arctic to the past 9000 years of global environmental change. New Phytologist 145, 257-269.
; Brincat et al., 2000Brincat, D., Yamada, K., Ishiwatari, R., Uemura, H., Naraoka, H. (2000) Molecular-isotopic stratigraphy of long-chain n-alkanes in Lake Baikal Holocene and glacial age sediments. Organic Geochemistry 31, 287-294.
; Rommerskirchen et al., 2006Rommerskirchen, F., Eglinton, G., Dupont, L., Rullkotter, J. (2006) Glacial/interglacial changes in southern Africa: Compound-specific δ13C land plant biomarker and pollen records from southeast Atlantic continental margin sediments. Geochemistry Geophysics Geosystems 7, doi: 10.1029/2005gc001223.
; Galy et al., 2008Galy, V., Francois, L., France-Lanord, C., Faure, P., Kudrass, H., Palhol, F., Singh, S.K. (2008) C4 plants decline in the Himalayan basin since the Last Glacial Maximum. Quaternary Science Reviews 27, 1396-1409.
; Norström et al., 2009Norström, E., Scott, L., Partridge, T.C., Risberg, J., Holmgren, K. (2009) Reconstruction of environmental and climate changes at Braamhoek wetland, eastern escarpment South Africa, during the last 16,000 years with emphasis on the Pleistocene-Holocene transition. Palaeogeography, Palaeoclimatology, Palaeoecology 271, 240-258.
; Pessenda et al., 2009Pessenda, L.C.R., De Oliveira, P.E., Mofatto, M., de Medeiros, V.B., Garcia, R.J.F., Aravena, R., Bendassoli, J.A., Leite, A.Z., Saad, A.R., Etchebehere, M.L. (2009) The evolution of a tropical rainforest/grassland mosaic in southeastern Brazil since 28,000 14C yr BP based on carbon isotopes and pollen records. Quaternary Research 71, 437-452.
; Sinninghe Damsté et al., 2011Sinninghe Damsté, J.S., Verschuren, D., Ossebaar, J., Blokker, J., van Houten, R., van der Meer, M.T.J., Plessen, B., Schouten, S. (2011) A 25,000-year record of climate-induced changes in lowland vegetation of eastern equatorial Africa revealed by the stable carbon-isotopic composition of fossil plant leaf waxes. Earth and Planetary Science Letters 302, 236-246.
; Lu et al., 2012Lu, H.Y., Zhou, Y.L., Liu, W.G., Mason, J. (2012) Organic stable carbon isotopic composition reveals late Quaternary vegetation changes in the dune fields of northern China. Quaternary Research 77, 433-444.
; Menking et al., 2012Menking, K.M., Peteet, D.M., Anderson, R.Y. (2012) Late-glacial and Holocene vegetation and climate variability, including major droughts, in the Sky Lakes region of southeastern New York State. Palaeogeography, Palaeoclimatology, Palaeoecology 353, 45-59.
; You and Liu, 2012You, H.T., Liu, J.Q. (2012) High-resolution climate evolution derived from the sediment records of Erlongwan Maar Lake since 14 ka BP. Chinese Science Bulletin 57, 3610-3616.
; Barker et al., 2013Barker, P.A., Hurrell, E.R., Leng, M.J., Plessen, B., Wolff, C., Conley, D.J., Keppens, E., Milne, I., Cumming, B.F., Laird, K.R., Kendrick, C.P., Wynn, P.M., Verschuren, D. (2013) Carbon cycling within an East African lake revealed by the carbon isotope composition of diatom silica: a 25-ka record from Lake Challa, Mt. Kilimanjaro. Quaternary Science Reviews 66, 55-63.
; Li et al., 2013Li, H.C., Liew, P.M., Seki, O., Kuo, T.S., Kawamura, K., Wang, L.C., Lee, T.Q. (2013) Paleoclimate variability in central Taiwan during the past 30 Kyrs reflected by pollen, δ13CTOC, and n-alkane-dD records in a peat sequence from Toushe Basin. Journal of Asian Earth Science 69, 166-176.
). Combining data for two studies of willow leaves in Sweden (Beerling, 1996Beerling, D.J. (1996) 13C discrimination by fossil leaves during the late-glacial climate oscillation 12-10 ka BP: Measurements and physiological controls. Oecologia 108, 29-37.
; Beerling and Rundgren, 2000Beerling, D.J., Rundgren, M. (2000) Leaf metabolic and morphological responses of dwarf willow (Salix herbacea) in the sub-arctic to the past 9000 years of global environmental change. New Phytologist 145, 257-269.
) and the parallel studies of Lake Challa (Sinninghe Damsté et al., 2011Sinninghe Damsté, J.S., Verschuren, D., Ossebaarvan der Sleen et al. (2015), J., Blokker, J., van Houten, R., van der Meer, M.T.J., Plessen, B., Schouten, S. (2011) A 25,000-year record of climate-induced changes in lowland vegetation of eastern equatorial Africa revealed by the stable carbon-isotopic composition of fossil plant leaf waxes. Earth and Planetary Science Letters 302, 236-246.
; Barker et al., 2013Barker, P.A., Hurrell, E.R., Leng, M.J., Plessen, B., Wolff, C., Conley, D.J., Keppens, E., Milne, I., Cumming, B.F., Laird, K.R., Kendrick, C.P., Wynn, P.M., Verschuren, D. (2013) Carbon cycling within an East African lake revealed by the carbon isotope composition of diatom silica: a 25-ka record from Lake Challa, Mt. Kilimanjaro. Quaternary Science Reviews 66, 55-63.
) yields 14 independent records (Fig. S-1), two of which were illustrated in Figure 2. Two datasets (You and Liu, 2012You, H.T., Liu, J.Q. (2012) High-resolution climate evolution derived from the sediment records of Erlongwan Maar Lake since 14 ka BP. Chinese Science Bulletin 57, 3610-3616.
; Li et al., 2013Li, H.C., Liew, P.M., Seki, O., Kuo, T.S., Kawamura, K., Wang, L.C., Lee, T.Q. (2013) Paleoclimate variability in central Taiwan during the past 30 Kyrs reflected by pollen, δ13CTOC, and n-alkane-dD records in a peat sequence from Toushe Basin. Journal of Asian Earth Science 69, 166-176.
) are quite scattered, so a 5-pt running average was used for plotting.These records more completely illustrate several principles discussed in the main text. Significantly, different ecosystem components can contribute carbon with very different δ13C and Δ13C values (Fig. S-1a). For example, average C3 biomass on Earth today (2000 AD) has a δ13C value of ~‑28.5 ‰ (Kohn, 2010
Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
), or Δ13C ~21.1 ‰. Soil organic matter typically has δ13C values ~2 ‰ higher than source leaves (Bowling et al., 2008Bowling, D.R., Pataki, D.E., Randerson, J.T. (2008) Carbon isotopes in terrestrial ecosystem pools and CO2 fluxes. New Phytologist 178, 24-40.
), implying Δ13C ~19.0 ‰. Extremely dry environments have the highest δ13C values for terrestrial C3 plants on Earth averaging ~‑24.5 ‰, implying a minimum C3 Δ13C(CO2-plant) of ~16.9 ‰ or a minimum C3-based Δ13C(CO2-SOM) ~14.8 ‰. Thus, Δ13C(CO2-SOM) < ~14 ‰ implies some carbon source other than pure C3 plants, e.g., C4 plants at low latitudes [Δ13C(CO2-C4) ~5 ‰]. At the other end of the isotopic spectrum, some lakes have abundant diatoms, whose δ13C values are quite low, e.g., c. ‑32 ‰ for Holocene Lake Challa (Barker et al., 2013Barker, P.A., Hurrell, E.R., Leng, M.J., Plessen, B., Wolff, C., Conley, D.J., Keppens, E., Milne, I., Cumming, B.F., Laird, K.R., Kendrick, C.P., Wynn, P.M., Verschuren, D. (2013) Carbon cycling within an East African lake revealed by the carbon isotope composition of diatom silica: a 25-ka record from Lake Challa, Mt. Kilimanjaro. Quaternary Science Reviews 66, 55-63.
), implying Δ13C(CO2-diatom) ≥ ~26 ‰.Some general features of these records stand out.
- (1) Clear advantages attend records derived from a single plant species (Fig. S-1b,c), as these are not subject to questions of C-source. Data scatter can be a problem, however, as illustrated for modern/historical tree rings (Fig. 2a).
- (2) Not all records are from a single locality (e.g., Fig. S-1b,g). While the necessity of splicing different data sources is completely understandable, it can raise questions about geographic variations, e.g., pine analyses for the Holocene in the western US tend to be from higher elevations and higher latitudes than for the Pleistocene (Fig. S-1b). Higher elevations in the western US tend to be wetter, which might explain some of the isotopic shift, and reduce any inferred dependence on pCO2.
- (3) Organic C preservation can potentially impact interpretations. The New York lake record (Fig. S-1d; Menking et al., 2012
Menking, K.M., Peteet, D.M., Anderson, R.Y. (2012) Late-glacial and Holocene vegetation and climate variability, including major droughts, in the Sky Lakes region of southeastern New York State. Palaeogeography, Palaeoclimatology, Palaeoecology 353, 45-59.
) shows a dramatic increase in Δ13C as C content increases from ~1 to 15 %. The British soil record shows a similar correlation between 2.5 and 40 % (Fig. S-1e; Huang et al., 1996Huang, Y.S., Bol, R., Harkness, D.D., Ineson, P., Eglinton, G. (1996) Post-glacial variations in distributions, C-13 and C-14 contents of aliphatic hydrocarbons and bulk organic matter in three types of British acid upland soils. Organic Geochemistry 24, 273-287.
). - (4) Low-latitude records are sensitive to changes in C4 biomass. Several studies interpreted isotopic shifts to reflect decreases in C4 abundances across the Pleistocene-Holocene transition (Fig. S-1f-j,o). C4 abundance is predicted to have been higher during the Pleistocene in many low-latitude areas because lower pCO2 stabilises C4 plants relative to C3 plants (Ehleringer et al., 1997
Ehleringer, J.R., Cerling, T.E., Helliker, B.R. (1997) C-4 photosynthesis, atmospheric CO2 and climate. Oecologia 112, 285-299.
). - (5) Changes to ecosystem vegetation in parallel with climate change can correlate with Δ13C (Fig. S-1k-m). Such correlations can occur because C3 plant δ13C depends on MAP (e.g., see Kohn, 2010
Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
), and different plant functional types exhibit different Δ13C (Diefendorf et al., 2010Diefendorf, A.F., Mueller, K.E., Wing, S.L., Koch, P.L., Freeman, K.H. (2010) Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proceedings of the National Academy of Sciences 107, 5738-5743.
). - (6) Diatoms can potentially impact δ13C of lake sediments. The shift in Δ13C for Lake Baikal (Fig. S-1k) corresponds with a dramatic change in the abundance of diatoms (Brincat et al., 2000
Brincat, D., Yamada, K., Ishiwatari, R., Uemura, H., Naraoka, H. (2000) Molecular-isotopic stratigraphy of long-chain n-alkanes in Lake Baikal Holocene and glacial age sediments. Organic Geochemistry 31, 287-294.
).
Figure S-1 (a) Principles of carbon isotope compositions relevant to interpreting isotopic records of sediment organic matter (SOM). C4 grasses have extremely low Δ13C, C3 plants have increasing Δ13C with increasing MAP, and diatoms have extremely high Δ13C. Applying a ~2 ‰ offset between plants and SOM (Bowling et al., 2008
Bowling, D.R., Pataki, D.E., Randerson, J.T. (2008) Carbon isotopes in terrestrial ecosystem pools and CO2 fluxes. New Phytologist 178, 24-40.
) implies a minimum cutoff of Δ13C(CO2-SOM) ~ 14 ‰ for which other sources of carbon besides C3 plants must be considered. (b-o) Records of Pleistocene-Holocene transition annotated with observations and interpretations from the original publications.The intent of these examples is not to argue that a pCO2-effect is unlikely, rather to emphasise that numerous other known factors must be quantitatively addressed before a pCO2-effect can be inferred from such records. Many records are susceptible to small changes in diatom and C4 plant abundance, as these have extreme Δ13C compared to terrestrial C3 plants.
SI-4: Why is Calculated MAP so Low in the Late Eocene and Oligocene?
The causes of relatively high δ13C(plant) (which lead to low calculated MAP) during late Eocene to early Oligocene are not immediately clear. Abundant high δ13C CAM or C4 plants seem unlikely. CAM rarely contributes significantly to ecosystem biomass or herbivore diets, and evidence for C4 plants prior to ~15 Ma remains meager and controversial. Certainly C4 plants did not constitute a significant proportion of large herbivore diets until the late Miocene (Cerling et al., 1997
Cerling, T.E., Harris, J.M., MacFadden, B.J., Leakey, M.G., Quade, J., Eisenmann, V., Ehleringer, J.R. (1997) Global vegetation change through the Miocene/Pliocene boundary. Nature 389, 153-158.
). If C4 plants were present, at least one taxon would probably consume it preferentially, yet δ13C values are not high enough to cross into a clear C4 isotopic field. Increases in water use efficiency in high pCO2 environments might allow floras (and herbivores) to survive in drier environments than commonly encountered on Earth today. Indeed a recent proxy for leaf area index indicates open environments in Patagonia through this interval (Dunn et al., 2015Dunn, R.E., Strömberg, C.A.E., Madden, R.H., Kohn, M.J., Carlini, A.A. (2015) Linked canopy, climate and faunal change in the Cenozoic of Patagonia. Science 347, 258-261.
), consistent with this view. Alternatively, δ13C values of atmospheric CO2 might be underestimated. Reevaluation of atmospheric δ13C calculations through the Cenozoic is beyond the scope of this study, but during the “dry” period, planktonic-benthic foraminifera Δ13C is unusually small (0-1 ‰) compared to all other times of the Cenozoic (1-2 ‰; Tipple et al., 2010Tipple, B.J., Meyers, S.R., Pagani, M. (2010) Carbon isotope ratio of Cenozoic CO2: a comparative evaluation of available geochemical proxies. Paleoceanography 25, doi: 10.1029/2009PA001851.
). This difference perhaps points to systematic differences in carbon cycle and ocean circulation during this time period, which might lead to systematic errors in calculated atmospheric δ13C. A 1 ‰ upward correction to atmospheric δ13C would increase MAP. For example, using data from Zanazzi and Kohn (2008Zanazzi, A., Kohn, M.J. (2008) Ecology and physiology of White River mammals based on stable isotope ratios of teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 257, 22-37.
; 35 Ma; pCO2 = 950 ppmv) the pCO2-dependent model MAP would increase from -200 to -100 mm/yr (still hyper-arid deserts), whereas the pCO2-independent model MAP would increase from ~200 to ~500 mm/yr. These changes would improve the correspondence between the MAP histogram and modern-day global distribution of MAP for the pCO2-independent model (Fig. 4b), but would not significantly affect the distribution of MAP for the pCO2-dependent model (Fig. 4c) because calculated MAP during this time interval would still be ≤0.Table S-3 Calculated pCO2-dependencies of δ13C in C3 plants.
| Record | pCO2 coefficient ±2 s.e. | pCO2 range |
| (‰/100 ppmv) | (ppmv, to nearest 5 ppmv) | |
| Modern leaves | -1.2 ± 1.0 | 325-380 |
| Tropical tree rings | 0.62 ± 1.05 | 285-390 |
| Borneo trees† | (0.0 ± 0.2) | 285-385 |
| European tree rings† | (-0.55 ± 0.67) | 300-370 |
| Sediment records† | (≤1.0 or ~0.0) | 185-280 |
| Pleistocene fossils* | -0.03 ± 0.11 | 190 to 370 |
| Paleogene fossils* | -0.03 ± 0.24 | 370 to 715 |
| Weighted average | -0.04 ± 0.26 | 190-715 |
†Not used in weighted average.
*Calculations are referenced to conditions at 2000 AD, which had pCO2 = 370 ppmv
Table S-4 Summary of records and factors influencing calculations.
| Data source | Largest uncertainties affecting calculations (magnitude, ‰) |
| Modern leaves | Data scatter (1-2 ‰) |
| Experiments | Poorly constrained MAP, plastic phenotypic response (unknown) |
| Tropical tree rings | Oscillations of unknown origin (1-2 ‰) |
| Pleistocene-Holocene sediments | MAP, C3/C4 changes, variable diatoms, variable C preservation, oscillations of unknown origin (2-5 ‰) |
| Quaternary fossils | Data scatter, uncertain tissue-diet fractionation (<1 ‰) |
| Tertiary fossils | Poorly constrained pCO2, MAP (<1 ‰) |
Supplementary Information References
Aiglstorfer, M., Bocherens, H., Bohme, M. (2014) Large mammal ecology in the late Middle Miocene Gratkorn locality (Austria). Palaeobiodiversity and Palaeoenvironments 94, 189-213.
Show in context Table S-2
View in article
Alder, J.R., Hostetler, S.W. (2015) Global climate simulations at 3000-year intervals for the last 21000 years with the GENMOM coupled atmosphere-ocean model. Climate of the Past 11, 449-471.
Show in context Independent MAP estimates were taken from regional floral studies (Wilf, 2000; Coltrain et al., 2004; Greenwood et al., 2010; Bartlein et al., 2011; Böhme et al., 2011; Melles et al., 2012), and/or from general circulation models (Henrot et al., 2010; Krapp and Jungclaus, 2011; Alder and Hostetler, 2015).
View in article
Floral studies (Bartlein et al., 2011) and the PMIP3 composite GCM results (Alder and Hostetler, 2015) both imply ΔMAP of ~-500 mm/yr during the Pleistocene, i.e. Pleistocene MAP = 650 mm/yr.
View in article
Balasse, M., Smith, A.B., Ambrose, S.H., Leigh, S.R. (2003) Determining sheep birth seasonality by analysis of tooth enamel oxygen isotopes ratios: the late Stone Age site of Kasteelberg (South Africa). Journal of Archaeological Science 30, 205-215.
Show in context Herbivore data represent highly integrated measures of leaf δ13C: if a 50 kg herbivore (e.g., sheep or goat), consumes c. 2 kg of leaves per day (Kohn, 1996), and each leaf weighs 0.5 g (c. 0.05 cm x 2 cm x 5 cm), a single bulk tooth enamel isotope composition that integrates 1-2 years of diet (Balasse et al., 2003) would represent approximately 2 million leaves.
View in article
Barker, P.A., Hurrell, E.R., Leng, M.J., Plessen, B., Wolff, C., Conley, D.J., Keppens, E., Milne, I., Cumming, B.F., Laird, K.R., Kendrick, C.P., Wynn, P.M., Verschuren, D.(2013) Carbon cycling within an East African lake revealed by the carbon isotope composition of diatom silica: a 25-ka record from Lake Challa, Mt. Kilimanjaro. Quaternary Science Reviews 66, 55-63.
Show in context Numerous records exist for sediment organic matter (SOM) spanning the Pleistocene-Holocene transition, but for simplicity only three localities were considered in detail: loessic sediments from the Rhine River valley in France and Germany (Fig. 2b; Hatté et al., 1998) and sediment cores from Lake Challa, Kenya (Fig. 2c; Sinninghe Damsté et al., 2011; Barker et al., 2013).
View in article
Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Combining data for two studies of willow leaves in Sweden (Beerling, 1996; Beerling and Rundgren, 2000) and the parallel studies of Lake Challa (Sinninghe Damsté et al., 2011; Barker et al., 2013) yields 14 independent records (Fig. S-1), two of which were illustrated in Figure 2.
View in article
At the other end of the isotopic spectrum, some lakes have abundant diatoms, whose δ13C values are quite low, e.g., c. ‑32 ‰ for Holocene Lake Challa (Barker et al., 2013), implying Δ13C(CO2-diatom) ≥ ~26 ‰.
View in article
Bartlein, P.J., Harrison, S.P., Brewer, S., Connor, S., Davis, B.A.S., Gajewski, K., Guiot, J., Harrison-Prentice, T.I., Henderson, A., Peyron, O., Prentice, I.C., Scholze, M., Seppa, H., Shuman, B., Sugita, S., Thompson, R.S., Viau, A.E., Williams, J., Wu, H. (2011) Pollen-based continental climate reconstructions at 6 and 21 ka: a global synthesis. Climate Dynamics 37, 775-802.
Show in context Independent MAP estimates were taken from regional floral studies (Wilf, 2000; Coltrain et al., 2004; Greenwood et al., 2010; Bartlein et al., 2011; Böhme et al., 2011; Melles et al., 2012), and/or from general circulation models (Henrot et al., 2010; Krapp and Jungclaus, 2011; Alder and Hostetler, 2015).
View in article
Floral studies (Bartlein et al., 2011) and the PMIP3 composite GCM results (Alder and Hostetler, 2015) both imply ΔMAP of ~-500 mm/yr during the Pleistocene, i.e. Pleistocene MAP = 650 mm/yr.
View in article
Beerling, D.J. (1996) 13C discrimination by fossil leaves during the late-glacial climate oscillation 12-10 ka BP: Measurements and physiological controls. Oecologia 108, 29-37.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Combining data for two studies of willow leaves in Sweden (Beerling, 1996; Beerling and Rundgren, 2000) and the parallel studies of Lake Challa (Sinninghe Damsté et al., 2011; Barker et al., 2013) yields 14 independent records (Fig. S-1), two of which were illustrated in Figure 2.
View in article
Beerling, D.J., Rundgren, M. (2000) Leaf metabolic and morphological responses of dwarf willow (Salix herbacea) in the sub-arctic to the past 9000 years of global environmental change. New Phytologist 145, 257-269.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Combining data for two studies of willow leaves in Sweden (Beerling, 1996; Beerling and Rundgren, 2000) and the parallel studies of Lake Challa (Sinninghe Damsté et al., 2011; Barker et al., 2013) yields 14 independent records (Fig. S-1), two of which were illustrated in Figure 2.
View in article
Beerling, D.J., Royer, D.L. (2011) Convergent Cenozoic CO2 history. Nature Geoscience 4, 418-420.
Show in context For data older than 3 Ma, pCO2 and δ13C values were estimated from geochemical proxies (Seki et al., 2010; Tipple et al., 2010; Beerling and Royer, 2011); the 3 Ma filtered estimates for δ13C of Tipple et al. (2010) were used, except for rapid changes across the Eocene-Oligocene transition, for which a more precise estimate was based on high-resolution foraminiferal records (Coxall et al., 2005).
View in article
Bereiter, B., Lüthi, D., Siegrist, M., Schupbach, S., Stocker, T.F., Fischer, H. (2012) Mode change of millennial CO2 variability during the last glacial cycle associated with a bipolar marine carbon seesaw. Proceedings of the National Academy of Sciences 109, 9755-9760.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25.
Show in context Table S-2
View in article
Biasatti, D., Wang, Y., Gao, F., Xu, Y., Flynn, L. (2012) Paleoecologies and paleoclimates of late Cenozoic mammals from Southwest China: evidence from stable carbon and oxygen isotopes. Journal of Asian Earth Science 44, 48-61.
Show in context Table S-2
View in article
Bibi, F. (2007) Dietary niche partitioning among fossil bovids in late Miocene C3 habitats: Consilience of functional morphology and stable isotope analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 253, 529-538.
Show in context Table S-2
View in article
Blondel, C., Bocherens, H., Mariotti, A. (1997) Stable carbon and oxygen isotope ratios in ungulate teeth from French Eocene and Oligocene localities. Bulletin Societé Geologique de France 168, 775-781.
Show in context Table S-2
View in article
Boardman, G.S., Secord, R. (2013) Stable isotope paleoecology of White River ungulates during the Eocene-Oligocene climate transition in northwestern Nebraska. Palaeogeography, Palaeoclimatology, Palaeoecology 375, 38-49.
Show in contextTable S-2
View in article
Bocherens, H., Drucker, D. (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from Recent and ancient terrestrial ecosystems. International Journal of Osteoarcheology 13, 46-53.
Show in context Table S-2
View in article
Bocherens, H., Drucker, D.G., Bonjean, D., Bridault, A., Conard, N.J., Cupillard, C., Germonpre, M., Honeisen, M., Munzel, S.C., Napierala, H., Patou-Mathis, M., Stephan, E., Uerpmann, H.P., Ziegler, R. (2011) Isotopic evidence for dietary ecology of cave lion (Panthera spelaea) in North-Western Europe: Prey choice, competition and implications for extinction. Quaternary International 245, 249-261.
Show in context Table S-2
View in article
Bocherens, H., Drucker, D.G., Germonpre, M., Laznickova-Galetova, M., Naito, Y.I., Wissing, C., Bruzek, J., Oliva, M. (2015a) Reconstruction of the Gravettian food-web at Predmosti I using multi-isotopic tracking (C-13, N-15, S-34) of bone collagen. Quaternary International 359, 211-228.
Show in context Table S-2
View in article
Bocherens, H., Hofman-Kaminska, E., Drucker, D.G., Schmolcke, U., Kowalczyk, R. (2015b) European Bison as a refugee species? Evidence from isotopic data on early Holocene Bison and other large herbivores in northern Europe. PLoS One 10, 10.1371/journal.pone.0115090.
Show in context Table S-2
View in article
Böhme, M., Winklhofer, M., Ilg A. (2011) Miocene precipitation in Europe: temporal trends and spatial gradients. Palaeogeography, Palaeoclimatology, Palaeoecology 304, 212-218.
Show in context Independent MAP estimates were taken from regional floral studies (Wilf, 2000; Coltrain et al., 2004; Greenwood et al., 2010; Bartlein et al., 2011; Böhme et al., 2011; Melles et al., 2012), and/or from general circulation models (Henrot et al., 2010; Krapp and Jungclaus, 2011; Alder and Hostetler, 2015).
View in article
Bowling, D.R., Pataki, D.E., Randerson, J.T. (2008) Carbon isotopes in terrestrial ecosystem pools and CO2 fluxes. New Phytologist 178, 24-40.
Show in context Soil organic matter typically has δ13C values ~2 ‰ higher than source leaves (Bowling et al., 2008), implying Δ13C ~ 19.0 ‰.
View in article
Figure S-1 [...] Applying a ~2 ‰ offset between plants and SOM (Bowling et al., 2008) implies a minimum cutoff of Δ13C(CO2-SOM) ~ 14 ‰ for which other sources of carbon besides C3 plants must be considered.
View in article
Brincat, D., Yamada, K., Ishiwatari, R., Uemura, H., Naraoka, H. (2000) Molecular-isotopic stratigraphy of long-chain n-alkanes in Lake Baikal Holocene and glacial age sediments. Organic Geochemistry 31, 287-294.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
(6) Diatoms can potentially impact δ13C of lake sediments. The shift in Δ13C for Lake Baikal (Fig. S-1k) corresponds with a dramatic change in the abundance of diatoms (Brincat et al., 2000).
View in article
Britton, K., Gaudzinski-Windheuser, S., Roebroeks, W., Kindler, L., Richards, M.P. (2012) Stable isotope analysis of well-preserved 120,000-year-old herbivore bone collagen from the Middle Palaeolithic site of Neumark-Nord 2, Germany reveals niche separation between bovids and equids. Palaeogeography, Palaeoclimatology, Palaeoecology 333, 168-177.
Show in context Table S-2
View in article
Castaños, J., Zuluaga, M.C., Ortega, L.A., Murelaga, X., Alonso-Olazabal, A., Rofes, J., Castaños, P. (2014) Carbon and nitrogen stable isotopes of bone collagen of large herbivores from the Late Pleistocene Kiputz IX cave site (Gipuzkoa, north Iberian Peninsula) for palaeoenvironmental reconstruction. Quaternary International 339, 131-138.
Show in context Table S-2
View in article
Cerling, T.E., Harris, J.M., MacFadden, B.J., Leakey, M.G., Quade, J., Eisenmann, V., Ehleringer, J.R. (1997) Global vegetation change through the Miocene/Pliocene boundary. Nature 389, 153-158.
Show in context Table S-2
View in article
Certainly C4 plants did not constitute a significant proportion of large herbivore diets until the late Miocene (Cerling et al., 1997).
View in article
Cerling, T.E., Harris, J.M. (1999) Carbon isotope fractionation between diet and bioapatite in ungulate mammals and implications for ecological and paleoecological studies. Oecologia 120, 347-363.
Show in context Herbivore-diet fractionations for tooth enamel assumed a 14 ‰ offset for perissodactyls (Cerling and Harris, 1999) and notoungulates (Kohn et al., 2015) and 14.5 ‰ for artiodactyls (Passey et al., 2005).
View in article
Coltrain, J.B., Harris, J.M., Cerling, T.E., Ehleringer, J.R., Dearing, M.-D., Ward, J., Allen, J. (2004) Rancho La Brea stable isotope biogeochemistry and its implications for the palaeoecology of late Pleistocene, coastal Southern California. Palaeogeography, Palaeoclimatology, Palaeoecology 205, 199-219.
Show in context Independent MAP estimates were taken from regional floral studies (Wilf, 2000; Coltrain et al., 2004; Greenwood et al., 2010; Bartlein et al., 2011; Böhme et al., 2011; Melles et al., 2012), and/or from general circulation models (Henrot et al., 2010; Krapp and Jungclaus, 2011; Alder and Hostetler, 2015).
View in article
Table S-2
View in article
Coxall, H.K., Wilson, P.A., Pälike, H., Lear, C.H., Backman, J. (2005) Rapid stepwise onset of Antarctic glaciation and deeper calcite compensation in the Pacific Ocean. Nature 433, 53-57.
Show in context For data older than 3 Ma, pCO2 and δ13C values were estimated from geochemical proxies (Seki et al., 2010; Tipple et al., 2010; Beerling and Royer, 2011); the 3 Ma filtered estimates for δ13C of Tipple et al. (2010) were used, except for rapid changes across the Eocene-Oligocene transition, for which a more precise estimate was based on high-resolution foraminiferal records (Coxall et al., 2005).
View in article
DeSantis, L.R.G., Wallace, S.C. (2008) Neogene forests from the Appalachians of Tennessee, USA: Geochemical evidence from fossil mammal teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 266, 59-68.
Show in context Table S-2
View in article
DeSantis, L.R.G., Feranec, R.S., MacFadden, B.J. (2009) Effects of Global Warming on Ancient Mammalian Communities and Their Environments. PLoS One 4, 10.1371/journal.pone.0005750.
Show in context Table S-2
View in article
Diefendorf, A.F., Mueller, K.E., Wing, S.L., Koch, P.L., Freeman, K.H. (2010) Global patterns in leaf 13C discrimination and implications for studies of past and future climate. Proceedings of the National Academy of Sciences 107, 5738-5743.
Show in context Such correlations can occur because C3 plant δ13C depends on MAP (g., see Kohn, 2010), and different plant functional types exhibit different Δ13C (Diefendorf et al., 2010).
View in article
Domingo, L., Koch, P.L., Grimes, S.T., Morales J., Lopez-Martinez N. (2012) Isotopic paleoecology of mammals and the Middle Miocene Cooling event in the Madrid Basin (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 339, 98-113.
Show in context Table S-2
View in article
Domingo, L., Koch, P.L., Fernandez, M.H., Fox, D.L., Domingo, M.S., Alberdi, M.T. (2013) Late Neogene and early Quaternary paleoenvironmental and paleoclimatic conditions in southwestern Europe: Isotopic analyses on mammalian taxa. PLoS One 8, doi: 10.1371/journal.pone.0063739.
Show in context Table S-2
View in article
Dunn, R.E., Strömberg, C.A.E., Madden, R.H., Kohn, M.J., Carlini, A.A. (2015) Linked canopy, climate and faunal change in the Cenozoic of Patagonia. Science 347, 258-261.
Show in context Indeed a recent proxy for leaf area index indicates open environments in Patagonia through this interval (Dunn et al., 2015), consistent with this view. Alternatively, δ13C values of atmospheric CO2 might be underestimated.
View in article
Eberle, J., Fricke, H., Humphrey, J. (2009) Lower-latitude mammals as year-round residents in Eocene Arctic forests. Geology 37, 499-502.
Show in context Table S-2
View in article
Ecker, M., Bocherens, H., Julien, M.A., Rivals, F., Raynal, J.P., Moncel, M.H. (2013) Middle Pleistocene ecology and Neanderthal subsistence: Insights from stable isotope analyses in Payre (Ardeche, southeastern France). Journal of Human Evolution 65, 363-373.
Show in contextTable S-2
View in article
Ehleringer, J.R., Cerling, T.E., Helliker, B.R. (1997) C-4 photosynthesis, atmospheric CO2 and climate. Oecologia 112, 285-299.
Show in contextC4 abundance is predicted to have been higher during the Pleistocene in many low-latitude areas because lower pCO2 stabilises C4 plants relative to C3 plants (Ehleringer et al., 1997).
View in article
Elsig, J., Schmitt, J., Leuenberger, D., Schneider, R., Eyer, M., Leuenberger, M., Joos, F., Fischer, H., Stocker, T.F. (2009) Stable isotope constraints on Holocene carbon cycle changes from an Antarctic ice core. Nature 461, 507-510.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
Farquhar, G.D., Ehleringer, J.R., Hubick, K.T. (1989) Carbon isotope discrimination and photosynthesis. Annual Review of Plant Physiology and Plant Molecular Biology 40, 503-537.
Show in context The slope for European trees was based on Figure 2a of Frank et al. (2015), specifically determining their best estimate of ci for years 1900 and 2000 AD, calculating a 2 standard error (s.e.) in ci from the spread of data, ratioing ci to known ca, and calculating Δ13C from standard expressions (Farquhar et al., 1989).
View in article
Feranec, R.S., MacFadden, B.J. (2006) Isotopic discrimination of resource partitioning among ungulates in C3-dominated communities from the Miocene of Florida and California. Paleobiology 32, 191-205.
Show in context Table S-2
View in article
Feranec, R.S., Garcia, N., Diez, J.C., Arsuaga, J.L. (2010) Understanding the ecology of mammalian carnivorans and herbivores from Valdegoba cave (Burgos, northern Spain) through stable isotope analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 263-272.
Show in context Table S-2
View in article
Fox-Dobbs, K., Leonard, J.A., Koch, P.L. (2008) Pleistocene megafauna from eastern Beringia: paleoecological and paleoenvironmental interpretations of stable carbon and nitrogen isotope and radiocarbon records. Palaeogeography, Palaeoclimatology, Palaeoecology 261, 30-46.
Show in context Data for Pleistocene caribou were omitted because consumption of high δ13C lichens biases their compositions relative to C3 consumers (Fox-Dobbs et al., 2008).
View in article
Table S-2
View in article
France, C.A.M., Zelanko, P.M., Kaufman, A.J., Holtz, T.R. (2007) Carbon and nitrogen isotopic analysis of Pleistocene mammals from the Saltville Quarry (Virginia, USA): Implications for trophic relationships. Palaeogeography, Palaeoclimatology, Palaeoecology 249, 271-282.
Show in context Table S-2
View in article
SI-2: Example Calculation – France et al. (2007)
View in article
Francey, R.J., Allison, C.E., Etheridge, D.M., Trudinger, C.M., Enting, I.G., Leuenberger, M., Langenfelds, R.L., Michel, E., Steele, L.P. (1999) A 1000-year high precision record of δ13C in atmospheric CO2. Tellus Series B 51, 170-193.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
Frank, D.C., Poulter, B., Saurer, M., Esper, J., Huntingford, C., Helle, G., Treydte, K., Zimmermann, N.E., Schleser, G.H., Ahlström, A., Ciais, P., Friedlingstein, P., Levis, S., Lomas, M., Sitch, S., Viovy, N., Andreu-Hayles, L., Bednarz, Z., Berninger, F., Boettger, T., D‘Alessandro, C.M., Daux, V., Filot, M., Grabner, M., Gutierrez, E., Haupt, M., Hilasvuori, E., Jungner, H., Kalela-Brundin, M., Krapiec, M., Leuenberger, M., Loader, N.J., Marah, H., Masson-Delmotte, V., Pazdur, A., Pawelczyk, S., Pierre, M., Planells, O., Pukiene, R., Reynolds-Henne, C.E., Rinne, K.T., Saracino, A., Sonninen, E., Stievenard, M., Switsur, V.R., Szczepanek, M., Szychowska-Krapiec, E., Todaro, L., Waterhouse, J.S., Weig, M. (2015) Water-use efficiency and transpiration across European forests during the Anthropocene. Nature Climate Change 5, 579–583, doi: 10.1038/NCLIMATE2614.
Show in context Results for individual tropical trees (Loader et al., 2011) and for European trees (Frank et al., 2015; their Fig. 2a) are also tabulated for reference, but were not used in calculating weighted mean values because of uncertainties in correcting for climate change (European dataset) and in accounting for ontogeny (individual tropical trees).
View in article
The slope for European trees was based on Figure 2a of Frank et al. (2015), specifically determining their best estimate of ci for years 1900 and 2000 AD, calculating a 2 standard error (s.e.) in ci from the spread of data, ratioing ci to known ca, and calculating Δ13C from standard expressions (Farquhar et al., 1989).
View in article
Table S-1 [...] European composite is from Frank et al. (2015) for Quercus and Pinus.
View in article
Froehle, A.W., Kellner, C.M., Schoeninger, M.J. (2010) FOCUS: effect of diet and protein source on carbon stable isotope ratios in collagen: follow up to Warinner and Tuross (2009). Journal of Archaeological Science 37, 2662-2670.
Show in context For collagen, a 5 ‰ offset was applied, relevant to wild herbivores consuming C3 plants, with an assumed uncertainty of ±0.5 ‰ (see summary of Froehle et al., 2010).
View in article
Galy, V., Francois, L., France-Lanord, C., Faure, P., Kudrass, H., Palhol, F., Singh, S.K. (2008) C4 plants decline in the Himalayan basin since the Last Glacial Maximum. Quaternary Science Reviews 27, 1396-1409.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Gilmour, D.M. (2011) Chronology and ecology of late Pleistocene megafauna in the northern Willamette Valley, Oregon. MS (Portland State University, Portland, Oregon).
Show in context Table S-2
View in article
Greenwood, D.R., Basinger, J.F., Smith, R.Y. (2010) How wet was the Arctic Eocene rain forest? Estimates of precipitation from Paleogene Arctic macrofloras. Geology 38, 15-18.
Show in contextIndependent MAP estimates were taken from regional floral studies (Wilf, 2000; Coltrain et al., 2004; Greenwood et al., 2010; Bartlein et al., 2011; Böhme et al., 2011; Melles et al., 2012), and/or from general circulation models (Henrot et al., 2010; Krapp and Jungclaus, 2011; Alder and Hostetler, 2015).
View in article
Hatté, C., Fontugne, M., Rousseau, D.D., Antoine, P., Zoller, L., Tisnerat-Laborde, N., Bentaleb, I. (1998) δ13C variations of loess organic matter as a record of the vegetation response to climatic changes during the Weichselian. Geology 26, 583-586.
Show in context Numerous records exist for sediment organic matter (SOM) spanning the Pleistocene-Holocene transition, but for simplicity only three localities were considered in detail: loessic sediments from the Rhine River valley in France and Germany (Fig. 2b; Hatté et al., 1998) and sediment cores from Lake Challa, Kenya (Fig. 2c; Sinninghe Damsté et al., 2011; Barker et al., 2013).
View in article
Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Henrot, A.J., François, L., Favre, E., Butzin, M., Ouberdous, M., Munhoven, G. (2010) Effects of CO2, continental distribution, topography and vegetation changes on the climate of the Middle Miocene: a model study. Climate of the Past 6, 675-694.
Show in context Independent MAP estimates were taken from regional floral studies (Wilf, 2000; Coltrain et al., 2004; Greenwood et al., 2010; Bartlein et al., 2011; Böhme et al., 2011; Melles et al., 2012), and/or from general circulation models (Henrot et al., 2010; Krapp and Jungclaus, 2011; Alder and Hostetler, 2015).
View in article
Huang, Y.S., Bol, R., Harkness, D.D., Ineson, P., Eglinton, G. (1996) Post-glacial variations in distributions, C-13 and C-14 contents of aliphatic hydrocarbons and bulk organic matter in three types of British acid upland soils. Organic Geochemistry 24, 273-287.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
The British soil record shows a similar correlation between 2.5 and 40 % (Fig. S-1e; Huang et al., 1996).
View in article
Iacumin, P., Bocherens, H., Delgado Huertas, A., Mariotti, A., Longinelli, A. (1997) A stable isotope study of fossil mammal remains from the Paglicci Cave, southern Italy: N and C as palaeoenvironmental indicators. Earth and Planetary Science Letters 148, 349-357.
Show in context Table S-2
View in article
Indermühle, A., Stocker, T.F., Joos, F., Fischer, H., Smith, H.J., Wahlen, M., Deck, B., Mastroianni, D., Tschumi, J., Blunier, T., Meyer, R., Stauffer, B. (1999) Holocene carbon-cycle dynamics based on CO2 trapped in ice at Taylor Dome, Antarctica. Nature 398, 121-126.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
Indermühle, A., Monnin, E., Stauffer, B., Stocker, T.F., Wahlen, M. (2000) Atmospheric CO2 concentration from 60 to 20 kyr BP from the Taylor Dome ice core, Antarctica. Geophysical Research Letters 27, 735-738.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
Koch, P.L., Zachos, J.C., Dettman, D.L. (1995) Stable isotope stratigraphy and paleoclimatology of the Paleogene Bighorn Basin (Wyoming, USA). Palaeogeography, Palaeoclimatology, Palaeoecology 115, 61-89.
Show in context Table S-2
View in article
Kocsis, L., Ozsvart, P., Becker, D., Ziegler, R., Scherler, L., Codrea, V. (2014) Orogeny forced terrestrial climate variation during the late Eocene-early Oligocene in Europe. Geology 42, 727-730.
Show in context Table S-2
View in article
Kohn, M.J. (1996) Predicting animal ä18O: Accounting for diet and physiological adaptation. Geochimica et Cosmochimica Acta 60, 4811-4829.
Show in context Herbivore data represent highly integrated measures of leaf δ13C: if a 50 kg herbivore (e.g., sheep or goat), consumes c. 2 kg of leaves per day (Kohn, 1996), and each leaf weighs 0.5 g (c. 0.05 cm x 2 cm x 5 cm), a single bulk tooth enamel isotope composition that integrates 1-2 years of diet (Balasse et al., 2003) would represent approximately 2 million leaves.
View in article
Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695.
Show in context For modern leaves, the dataset of Kohn (2010) was used, adding values for modern changes to pCO2 as determined from recent compilations (www.esrl.noaa.gov/gmd/ccgg/trends/; http://cdiac.esd.ornl.gov/trends/trends.htm; McCarroll and Loader, 2004) as an independent variable.
View in article
This dataset omits studies with a suspected C4 bias, represents a substantial increase over the 22 studies discussed in Kohn (2010), and particularly emphasises low-pCO2 (Pleistocene) and high-pCO2 (mid-Miocene and Eocene) times in the context of independently estimated MAP, pCO2, and the δ13C of atmospheric CO2.
View in article
Table S-2
View in article
Calculated Δ13C for the pCO2-independent model was based on equation 2 of Kohn (2010) using MAP, altitude and latitude as input parameters.
View in article
Isotopically-based MAP (Figs. 3c,d, 4) used the model of Kohn (2010), either directly (pCO2-independent) or after first correcting compositions for potential pCO2-dependence (Schubert and Jahren, 2012) based on independently measured or estimated pCO2.
View in article
For the site’s altitude (c. 500 m) and latitude (37 °N), equation 2 of Kohn (2010) yields an apparent MAP of 584 mm/yr.
View in article
Using this value as independent input to equation 2 of Kohn (2010), an expected Δ13C value of 19.9 ‰ was calculated for this latitude and altitude.
View in article
For example, average C3 biomass on Earth today (2000 AD) has a δ13C value of ~‑28.5 ‰ (Kohn, 2010), or Δ13C ~ 21.1 ‰.
View in article
Such correlations can occur because C3 plant δ13C depends on MAP (g., see Kohn, 2010), and different plant functional types exhibit different Δ13C (Diefendorf et al., 2010).
View in article
Kohn, M.J., McKay, M.P. (2012) Paleoecology of late Pleistocene-Holocene faunas of eastern and central Wyoming, USA, with implications for LGM climate models. Palaeogeography, Palaeoclimatology, Palaeoecology 326-328, 42-53.
Show in context Table S-2
View in article
Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37.
Show in context Table S-2
View in article
Herbivore-diet fractionations for tooth enamel assumed a 14 ‰ offset for perissodactyls (Cerling and Harris, 1999) and notoungulates (Kohn et al., 2015) and 14.5 ‰ for artiodactyls (Passey et al., 2005).
View in article
Krapp, M., Jungclaus, J.H. (2011) The Middle Miocene climate as modelled in an atmosphere-ocean-biosphere model. Climate of the Past 7, 1169-1188.
Show in context Independent MAP estimates were taken from regional floral studies (Wilf, 2000; Coltrain et al., 2004; Greenwood et al., 2010; Bartlein et al., 2011; Böhme et al., 2011; Melles et al., 2012), and/or from general circulation models (Henrot et al., 2010; Krapp and Jungclaus, 2011; Alder and Hostetler, 2015).
View in article
Kuitems, M., van der Plicht, J., Druckerc, D.G., van Kolfschotena, T., Palstrab, S.W.L., Bocherens, H. (2015) Carbon and nitrogen stable isotopes of well-preserved Middle Pleistocene bone collagen from Schöningen (Germany) and their palaeoecological implications. Journal of Human Evolution, doi: 10.1016/j.jhevol.2015.1001.1008.
Show in context Table S-2
View in article
Li, H.C., Liew, P.M., Seki, O., Kuo, T.S., Kawamura, K., Wang, L.C., Lee, T.Q. (2013) Paleoclimate variability in central Taiwan during the past 30 Kyrs reflected by pollen, δ13CTOC, and n-alkane-δD records in a peat sequence from Toushe Basin. Journal of Asian Earth Science 69, 166-176.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Two datasets (You and Liu, 2012; Li et al., 2013) are quite scattered, so a 5-pt running average was used for plotting.
View in article
Loader, N.J., Walsh, R.P.D., Robertson, I., Bidin, K., Ong, R.C., Reynolds, G., McCarroll, D., Gagen, M., Young, G.H.F. (2011) Recent trends in the intrinsic water-use efficiency of ringless rainforest trees in Borneo. Philosophical Transactions of the Royal Society B-Biological Science 366, 3330-3339.
Show in context Results for individual tropical trees (Loader et al., 2011) and for European trees (Frank et al., 2015; their Fig. 2a) are also tabulated for reference, but were not used in calculating weighted mean values because of uncertainties in correcting for climate change (European dataset) and in accounting for ontogeny (individual tropical trees).
View in article
Data from Loader et al. (2011) were regressed similarly to van der Sleen et al. (2015) – Δ13C values were calculated from tabulated data and regressed vs. pCO2.
View in article
Table S-1 *Regressions for specific tree ring data from van der Sleen et al. (2015) (Thailand, Cameroon, Bolivia; multiple trees) and Loader et al. (2011; Borneo; single trees).
View in article
Lourantou, A., Lavric, J.V., Kohler, P., Barnola, J.M., Paillard, D., Michel, E., Raynaud, D., Chappellaz, J. (2010) Constraint of the CO2 rise by new atmospheric carbon isotopic measurements during the last deglaciation. Global Biogeochemical Cycles 24, doi: 10.1029/2009gb003545.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
For the age of the site (16 ka) an atmospheric δ13C value of -6.8 ‰ was estimated from ice core records (Smith et al., 1999; Lourantou et al., 2010), yielding Δ13C (CO2-plant) = 19.7 ‰.
View in article
Lu, H.Y., Zhou, Y.L., Liu, W.G., Mason, J. (2012) Organic stable carbon isotopic composition reveals late Quaternary vegetation changes in the dune fields of northern China. Quaternary Research 77, 433-444.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Lüthi, D., Le Floch, M., Bereiter, B., Blunier, T., Barnola, J.M., Siegenthaler, U., Raynaud, D., Jouzel, J., Fischer, H., Kawamura, K., Stocker, T.F. (2008) High-resolution carbon dioxide concentration record 650,000-800,000 years before present. Nature 453, 379-382.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
MacFadden, B.J., Higgins, P. (2004) Ancient ecology of 15-million-year-old browsing mammals within C3 plant communities from Panama. Oecologia 140, 169-182.
Show in context Table S-2
View in article
Martin, C., Bentaleb, I., Antoine, P.O. (2011) Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 19-29.
Show in context Table S-2
View in article
Maung-Thein, Z.-M. Takai, M., Uno, H., Wynn, J.G., Egi, N., Tsubamoto, T., Thaung-Htike, Aung-Naing-Soe, Maung-Maung, Nishimura, T., Yoneda, M. (2011) Stable isotope analysis of the tooth enamel of Chaingzauk mammalian fauna (late Neogene, Myanmar) and its implication to paleoenvironment and paleogeography. Palaeogeography, Palaeoclimatology, Palaeoecology 300, 11-22.
Show in context Table S-2
View in article
McCarroll, D., Loader, N.J. (2004) Stable isotopes in tree rings. Quaternary Science Reviews 23, 771-801.
Show in context For modern leaves, the dataset of Kohn (2010) was used, adding values for modern changes to pCO2 as determined from recent compilations (www.esrl.noaa.gov/gmd/ccgg/trends/; http://cdiac.esd.ornl.gov/trends/trends.htm; McCarroll and Loader, 2004) as an independent variable.
View in article
For modern data (last few centuries), pCO2 and δ13C are commonly provided in the supplemental files of the original publications, and are based on direct measures of atmospheric CO2 and its isotope composition (McCarroll and Loader, 2004; www.esrl.noaa.gov/gmd/ccgg/trends/; http://cdiac.esd.ornl.gov/trends/trends.htm).
View in article
Medina, E., Minchin, P. (1980) Stratification of δ13C values of leaves in Amazonian rain forests. Oecologia 45, 377-378.
Show in context For tropical trees, data from van der Sleen et al. (2015) for 27 cm-diameter trees (canopy) were emphasised rather than 8 cm-diameter tree data (understory), because isotopes in closed canopy forest understory can depend on factors such as light intensity, humidity gradients, accumulation of ground-level CO2, and recycling of soil carbon (e.g., Vogel, 1978; Medina and Minchin, 1980; Ometto et al., 2006).
View in article
Melles, M., Brigham-Grette, J., Minyuk, P.S., Nowaczyk, N.R., Wennrich, V., DeConto, R.M., Anderson, P.M., Andreev, A.A., Coletti, A., Cook, T.L., Haltia-Hovi, E., Kukkonen, M., Lozhkin, A.V., Rosen, P., Tarasov, P., Vogel, H., Wagner, B. (2012) 2.8 Million Years of Arctic Climate Change from Lake El'gygytgyn, NE Russia. Science 337, 315-320.
Show in context Independent MAP estimates were taken from regional floral studies (Wilf, 2000; Coltrain et al., 2004; Greenwood et al., 2010; Bartlein et al., 2011; Böhme et al., 2011; Melles et al., 2012), and/or from general circulation models (Henrot et al., 2010; Krapp and Jungclaus, 2011; Alder and Hostetler, 2015).
View in article
Menking, K.M., Peteet, D.M., Anderson, R.Y. (2012) Late-glacial and Holocene vegetation and climate variability, including major droughts, in the Sky Lakes region of southeastern New York State. Palaeogeography, Palaeoclimatology, Palaeoecology 353, 45-59.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
The New York lake record (Fig. S-1d; Menking et al., 2012) shows a dramatic increase in Δ13C as C content increases from ~1 to 15 %.
View in article
Merceron, G., Zazzo, A., Spassov, N., Geraads, D., Kovachev, D. (2006) Bovid paleoecology and paleoenvironments from the Late Miocene of Bulgaria: evidence from dental microwear and stable isotopes. Palaeogeography, Palaeoclimatology, Palaeoecology 241, 637-654.
Show in context Table S-2
View in article
Montanari, S., Louys, J., Price, G.J. (2013) Pliocene paleoenvironments of southeastern Queensland, Australia inferred from stable isotopes of marsupial tooth enamel. PLoS One 8, doi: 10.1371/journal.pone.0066221.
Show in context Table S-2
View in article
Morgan, M.E., Kingston, J.D., Marino, B.D. (1994) Carbon isotopic evidence for the emergence of C4 plants in the Neogene from Pakistan and Kenya. Nature 367, 162-165.
Show in context Table S-2
View in article
Nelson, S.V. (2007) Isotopic reconstructions of habitat change surrounding the extinction of Sivapithecus, a Miocene hominoid, in the Siwalik Group of Pakistan. Palaeogeography, Palaeoclimatology, Palaeoecology 243, 204-222.
Show in context Table S-2
View in article
Norström, E., Scott, L., Partridge, T.C., Risberg, J., Holmgren, K. (2009) Reconstruction of environmental and climate changes at Braamhoek wetland, eastern escarpment South Africa, during the last 16,000 years with emphasis on the Pleistocene-Holocene transition. Palaeogeography, Palaeoclimatology, Palaeoecology 271, 240-258.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Ometto, J.P.H.B., Ehleringer, J.R., Domingues, T.F., Berry, J.A., I., F.Y., Mazzi, E., Higuchi, N., Flanagan, L.B., Nardoto, G.B., Martinelli, L.A. (2006) The stable carbon and nitrogen isotopic composition of vegetation in tropic forests of the Amazon Basin, Brazil. Biogeochemistry 79, 251-274.
Show in context For tropical trees, data from van der Sleen et al. (2015) for 27 cm-diameter trees (canopy) were emphasised rather than 8 cm-diameter tree data (understory), because isotopes in closed canopy forest understory can depend on factors such as light intensity, humidity gradients, accumulation of ground-level CO2, and recycling of soil carbon (e.g., Vogel, 1978; Medina and Minchin, 1980; Ometto et al., 2006).
View in article
Passey, B.H., Cerling, T.E., Perkins, M.E., Voorhies, M.R., Harris, J.M., Tucker, S.T. (2002) Environmental change in the Great Plains; an isotopic record from fossil horses. Journal of Geology 110, 123-140.
Show in context Table S-2
View in article
Passey, B.H., Robinson, T.F., Ayliffe, L.K., Cerling, T.E., Sponheimer, M., Dearing, M.D., Roeder, B.L., Ehleringer, J.R. (2005) Carbon isotope fractionation between diet, breath CO2, and bioapatite in different mammals. Journal of Archaeological Science 32, 1459-1470.
Show in context Herbivore-diet fractionations for tooth enamel assumed a 14 ‰ offset for perissodactyls (Cerling and Harris, 1999) and notoungulates (Kohn et al., 2015) and 14.5 ‰ for artiodactyls (Passey et al., 2005).
View in article
Pedro, J.B., Rasmussen, S.O., van Ommen, T.D. (2012) Tightened constraints on the time-lag between Antarctic temperature and CO2 during the last deglaciation. Climate of the Past 8, 1213-1221.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
Application of equation 6 of Schubert and Jahren (2012), accounting for lower pCO2 (230 ppmv; Pedro et al., 2012) relative to reference conditions (AD 2000, pCO2 = 370 ppmv) implies corrected Δ13C = 22.3 ‰, or MAP = 2130 mm/yr.
View in article
Pessenda, L.C.R., De Oliveira, P.E., Mofatto, M., de Medeiros, V.B., Garcia, R.J.F., Aravena, R., Bendassoli, J.A., Leite, A.Z., Saad, A.R., Etchebehere, M.L. (2009) The evolution of a tropical rainforest/grassland mosaic in southeastern Brazil since 28,000 14C yr BP based on carbon isotopes and pollen records. Quaternary Research 71, 437-452.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Petit, J.R., Jouzel, J., Raynaud, D., Barkov, N.I., Barnola, J.M., Basile, I., Bender, M., Chappellaz, J., Davis, M., Delaygue, G., Delmotte, M., Kotlyakov, V.M., Legrand, M., Lipenkov, V.Y., Lorius, C., Pepin, L., Ritz, C., Saltzman, E., Stievenard, M. (1999) Climate and atmospheric history of the past 420,000 years from the Vostok ice core, Antarctica. Nature 399, 429-436.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
Pushkina, D., Bocherens, H., Ziegler, R. (2014) Unexpected palaeoecological features of the Middle and Late Pleistocene large herbivores in southwestern Germany revealed by stable isotopic abundances in tooth enamel. Quaternary International 339, 164-178.
Show in context Table S-2
View in article
Qu, Y.T., Jin, C.Z., Zhang, Y.Q., Hu, Y.W., Shang, X., Wang, C.S. (2014) Preservation assessments and carbon and oxygen isotopes analysis of tooth enamel of Gigantopithecus blacki and contemporary animals from Sanhe Cave, Chongzuo, South China during the Early Pleistocene. Quaternary International 354, 52-58.
Show in context Table S-2
View in article
Raghavan, M., Themudo, G.E., Smith, C.I., Zazula, G., Campos, P.F. (2014) Musk ox (Ovibos moschatus) of the mammoth steppe: tracing palaeodietary and palaeoenvironmental changes over the last 50,000 years using carbon and nitrogen isotopic analysis. Quaternary Science Reviews 102, 192-201.
Show in context Table S-2
View in article
Richards, M.P., Hedges, R.E.M. (2003) Variations in bone collagen δ13C and d15N values of fauna from Northwest Europe over the last 40000 years. Palaeogeography, Palaeoclimatology, Palaeoecology 193, 261-267.
Show in context Table S-2
View in article
Rommerskirchen, F., Eglinton, G., Dupont, L., Rullkotter, J. (2006) Glacial/interglacial changes in southern Africa: Compound-specific δ13C land plant biomarker and pollen records from southeast Atlantic continental margin sediments. Geochemistry Geophysics Geosystems 7, doi: 10.1029/2005gc001223.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Scherler, L., Tütken, T., Becker, D. (2014) Carbon and oxygen stable isotope compositions of late Pleistocene mammal teeth from dolines of Ajoie (Northwestern Switzerland). Quaternary Research 82, 378-387.
Show in context Table S-2
View in article
Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43.
Show in context Figure 2a was constructed by comparing values for Δ13C(CO2-plant) to a pCO2-independent model (constant Δ), the hyperbolic pCO2-dependent model (c. 1.5 ‰/100 ppmv between 280 and 390 ppmv; Schubert and Jahren, 2012), and, for reference, a constant intermediate pCO2-dependence (0.75 ‰/100 ppmv).
View in article
Table S-2 [...] S = model of Schubert and Jahren (2012) (pCO2-dependent)
View in article
For the pCO2-dependent model, these Δ13C values were then corrected according to Schubert and Jahren (2012).
View in article
Isotopically-based MAP (Figs. 3c,d, 4) used the model of Kohn (2010), either directly (pCO2-independent) or after first correcting compositions for potential pCO2-dependence (Schubert and Jahren, 2012) based on independently measured or estimated pCO2.
View in article
Application of equation 6 of Schubert and Jahren (2012), accounting for lower pCO2 (230 ppmv; Pedro et al., 2012) relative to reference conditions (AD 2000, pCO2 = 370 ppmv) implies corrected Δ13C = 22.3 ‰, or MAP = 2130 mm/yr.
View in article
Equation 6 of Schubert and Jahren (2012) implies an additional ~2.6 ‰ correction, yielding an expected Δ13C value of 22.5 ‰. The value determined from isotope measurements is 19.7 ‰.
View in article
Schubert, BA., Jahren, A.H. (2015) Global increase in plant carbon isotope fractionation following the Last Glacial Maximum caused by increase in atmospheric pCO2. Geology 43, 435-438.
Show in context These records were chosen as characteristic of mid-latitude and low-latitude sites, and exemplify both the increase in Δ13C(CO2-SOM) commonly observed between c. 20 and c. 10 ka (Schubert and Jahren, 2015), and the issues involved in interpreting these isotopic shifts.
View in article
Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Secord, R., Wing, S.L., Chew, A. (2008) Stable isotopes in early Eocene mammals as indicators of forest canopy structure and resource partitioning. Paleobiology 34, 282-300.
Show in context Table S-2
View in article
Secord, R., Gingerich, P.D., Lohmann, K.C., MacLeod, K.G. (2010) Continental warming preceding the Palaeocene-Eocene thermal maximum. Nature 467, 955-958.
Show in context Table S-2
View in article
Ségalen, L., Lee-Thorp, J.A. (2009) Palaeoecology of late Early Miocene fauna in the Namib based on 13C/12C and 18O/16O ratios of tooth enamel and ratite eggshell carbonate. Palaeogeography, Palaeoclimatology, Palaeoecology 277, 191-198.
Show in context Table S-2
View in article
Seki, O., Foster, G.L., Schmidt, D.N., Mackensen, A., Kawamura, K., Pancost, R.D. (2010) Alkenone and boron-based Pliocene pCO2 records. Earth and Planetary Science Letters 292, 201-211.
Show in context For data older than 3 Ma, pCO2 and δ13C values were estimated from geochemical proxies (Seki et al., 2010; Tipple et al., 2010; Beerling and Royer, 2011); the 3 Ma filtered estimates for δ13C of Tipple et al. (2010) were used, except for rapid changes across the Eocene-Oligocene transition, for which a more precise estimate was based on high-resolution foraminiferal records (Coxall et al., 2005).
View in article
Siegenthaler, U., Stocker, T.F., Monnin, E., Lüthi, D., Schwander, J., Stauffer, B., Raynaud, D., Barnola, J.M., Fischer, H., Masson-Delmotte, V., Jouzel, J. (2005) Stable carbon cycle-climate relationship during the late Pleistocene. Science 310, 1313-1317.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
Sinninghe Damsté, J.S., Verschuren, D., Ossebaar, J., Blokker, J., van Houten, R., van der Meer, M.T.J., Plessen, B., Schouten, S. (2011) A 25,000-year record of climate-induced changes in lowland vegetation of eastern equatorial Africa revealed by the stable carbon-isotopic composition of fossil plant leaf waxes. Earth and Planetary Science Letters 302, 236-246.
Show in context Numerous records exist for sediment organic matter (SOM) spanning the Pleistocene-Holocene transition, but for simplicity only three localities were considered in detail: loessic sediments from the Rhine River valley in France and Germany (Fig. 2b; Hatté et al., 1998) and sediment cores from Lake Challa, Kenya (Fig. 2c; Sinninghe Damsté et al., 2011; Barker et al., 2013).
View in article
Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Combining data for two studies of willow leaves in Sweden (Beerling, 1996; Beerling and Rundgren, 2000) and the parallel studies of Lake Challa (Sinninghe Damsté et al., 2011; Barker et al., 2013) yields 14 independent records (Fig. S-1), two of which were illustrated in Figure 2.
View in article
Smith, H.J., Fischer, H., Wahlen, M., Mastroianni, D., Deck, B. (1999) Dual modes of the carbon cycle since the Last Glacial Maximum. Nature 400, 248-250.
Show in context For Pleistocene data, ice-core records were spliced and average pCO2 and δ13C values were estimated at 1000 year intervals (data from Francey et al., 1999; Indermühle et al., 1999, 2000; Smith et al., 1999; Siegenthaler et al., 2005; Lüthi et al., 2008; Elsig et al., 2009; Petit et al., 1999; Lourantou et al., 2010; Bereiter et al., 2012; Pedro et al., 2012).
View in article
For the age of the site (16 ka) an atmospheric δ13C value of -6.8 ‰ was estimated from ice core records (Smith et al., 1999; Lourantou et al., 2010), yielding Δ13C (CO2-plant) = 19.7 ‰.
View in article
Suraprasit, K., Chaimanee, Y., Bocherens, H., Chavasseau, O., Jaeger, J.J. (2014) Systematics and phylogeny of middle Miocene Cervidae (Mammalia) from Mae Moh Basin (Thailand) and a paleoenvironmental estimate using enamel isotopy of sympatric herbivore species. Journal of Vertebrate Paleontology 34, 179-194.
Show in context Table S-2
View in article
Tipple, B.J., Meyers, S.R., Pagani, M. (2010) Carbon isotope ratio of Cenozoic CO2: a comparative evaluation of available geochemical proxies. Paleoceanography 25, doi: 10.1029/2009PA001851.
Show in context Table S-2 [...] Note: Uncertainties in δ13C of atmospheric CO2 for the pre-Pleistocene is ~±0.35 ‰ (Tipple et al., 2010).
View in article
For data older than 3 Ma, pCO2 and δ13C values were estimated from geochemical proxies (Seki et al., 2010; Tipple et al., 2010; Beerling and Royer, 2011); the 3 Ma filtered estimates for δ13C of Tipple et al. (2010) were used, except for rapid changes across the Eocene-Oligocene transition, for which a more precise estimate was based on high-resolution foraminiferal records (Coxall et al., 2005).
View in article
Errors in δ13C were assumed to be ≤0.1 ‰ for the Pleistocene, and ±0.35 ‰ for older materials (Tipple et al., 2010).
View in article
Reevaluation of atmospheric δ13C calculations through the Cenozoic is beyond the scope of this study, but during the “dry” period, planktonic-benthic foraminifera Δ13C is unusually small (0-1 ‰) compared to all other times of the Cenozoic (1-2 ‰; Tipple et al., 2010).
View in article
Tütken, T., Vennemann, T. (2009) Stable isotope ecology of Miocene large mammals from Sandelzhausen, southern Germany. Paläontologische Zeitschrift 83, 207-226.
Show in context Table S-2
View in article
Tütken, T., Vennemann, T.W., Janz, H., Heizmann, E.P.J. (2006) Palaeoenvironment and palaeoclimate of the Middle Miocene lake in the Steinheim basin, SW Germany: a reconstruction from C, O, and Sr isotopes of fossil remains. Palaeogeography, Palaeoclimatology, Palaeoecology 241, 457-491.
Show in context Table S-2
View in article
Tütken, T., Kaiser, T.M., Vennemann, T., Merceron, G. (2013) Opportunistic feeding strategy for the earliest Old World hypsodont equids: Evidence from stable isotope and dental wear proxies. PLoS One 8, doi: 10.1371/journal.pone.0074463.
Show in contextTable S-2
View in article
Uno, K.T., Cerling, T.E., Harris, J.M., Kunimatsu, Y., Leakey, M.G., Nakatsukasa, M., Nakaya, H. (2011) Late Miocene to Pliocene carbon isotope record of differential diet change among East African herbivores. Proceedings of the National Academy of Sciences 108, 6509-6514.
Show in context Table S-2
View in article
van Dam, J.A., Reichart, G.J. (2009) Oxygen and carbon isotope signatures in late Neogene horse teeth from Spain and application as temperature and seasonality proxies. Palaeogeography, Palaeoclimatology, Palaeoecology 274, 64-81.
Show in context Table S-2
View in article
Van de Water, P.K., Leavitt, S.W., Betancourt, J.L. (1994) Trends in stomatal density and 13C/12C ratios of Pinus flexilis needles during last Glacial-Interglacial cycle. Science 264, 239-243.
Show in contextRecords that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
van der Sleen, P., Groenendijk, P., Vlam, M., Anten, N.P.R., Boom, A., Bongers, F., Pons, T.L., Terburg, G., Zuidema, P.A. (2015) No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience 8, 24-28.
Show in context For tropical trees, data from van der Sleen et al. (2015) for 27 cm-diameter trees (canopy) were emphasised rather than 8 cm-diameter tree data (understory), because isotopes in closed canopy forest understory can depend on factors such as light intensity, humidity gradients, accumulation of ground-level CO2, and recycling of soil carbon (e.g., Vogel, 1978; Medina and Minchin, 1980; Ometto et al., 2006).
View in article
Isotopic data were ordered according to age and values for Δ13C(CO2-plant) were calculated using records of atmospheric δ13CCO2 as tabulated in the supplemental files of van der Sleen et al. (2015).
View in article
Data from Loader et al. (2011) were regressed similarly to van der Sleen et al. (2015) – Δ13C values were calculated from tabulated data and regressed vs. pCO2.
View in article
Table S-1 *Regressions for specific tree ring data from van der Sleen et al. (2015) (Thailand, Cameroon, Bolivia; multiple trees) and Loader et al. (2011; Borneo; single trees).
View in article
Vogel, J.C. (1978) Recycling of carbon in a forest environment. Oecologia Plantarum 13, 89-94.
Show in context For tropical trees, data from van der Sleen et al. (2015) for 27 cm-diameter trees (canopy) were emphasised rather than 8 cm-diameter tree data (understory), because isotopes in closed canopy forest understory can depend on factors such as light intensity, humidity gradients, accumulation of ground-level CO2, and recycling of soil carbon (e.g., Vogel, 1978; Medina and Minchin, 1980; Ometto et al., 2006).
View in article
Wang, Y., Deng, T. (2005) A 25 m.y. isotopic record of paleodiet and environmental change from fossil mammals and paleosols from the NE margin of the Tibetan Plateau. Earth and Planetary Science Letters 236, 322-338.
Show in context Table S-2
View in article
White, T.D., Ambrose, S.H., Suwa, G., Su, D.F., DeGusta, D., Bernor, R.L., Boisserie, J.-R., Brunet, M., Delson, E., Frost, S., Garcia, N., Giaourtsakis, I.X., Haile-Selassie, Y., Howell, F.C., Lehmann, T., Likius, A., Pehlevan, C., Saegusa, H., Semprebon, G., Teaford, M., Vrba, E. (2009) Macrovertebrate paleontology and the Pliocene habitat of Ardipithecus ramidus. Science 326, 87-93.
Show in context Table S-2
View in article
Wilf, P. (2000) Late Paleocene-early Eocene climate changes in southwestern Wyoming: paleobotanical analysis. Geological Society of America Bulletin 112, 292-307.
Show in context Independent MAP estimates were taken from regional floral studies (Wilf, 2000; Coltrain et al., 2004; Greenwood et al., 2010; Bartlein et al., 2011; Böhme et al., 2011; Melles et al., 2012), and/or from general circulation models (Henrot et al., 2010; Krapp and Jungclaus, 2011; Alder and Hostetler, 2015).
View in article
You, H.T., Liu, J.Q. (2012) High-resolution climate evolution derived from the sediment records of Erlongwan Maar Lake since 14 ka BP. Chinese Science Bulletin 57, 3610-3616.
Show in context Records that cross the Pleistocene-Holocene transition (leaves and sediments) were extracted from the summary of Schubert and Jahren (2015), but were limited to only the 16 studies that provided at least 10 data points total, with at least 3 analyses prior to 10 ka and 3 analyses after 10 ka (Van de Water et al., 1994; Beerling, 1996; Huang et al., 1996; Hatté et al., 1998; Beerling and Rundgren, 2000; Brincat et al., 2000; Rommerskirchen et al., 2006; Galy et al., 2008; Norström et al., 2009; Pessenda et al., 2009; Sinninghe Damsté et al., 2011; Lu et al., 2012; Menking et al., 2012; You and Liu, 2012; Barker et al., 2013; Li et al., 2013).
View in article
Two datasets (You and Liu, 2012; Li et al., 2013) are quite scattered, so a 5-pt running average was used for plotting.
View in article
Zanazzi, A., Kohn, M.J. (2008) Ecology and physiology of White River mammals based on stable isotope ratios of teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 257, 22-37.
Show in context Table S-2
View in article
For example, using data from Zanazzi and Kohn (2008; 35 Ma; pCO2 = 950 ppmv) the pCO2-dependent model MAP would increase from -200 to -100 mm/yr (still hyper-arid deserts), whereas the pCO2-independent model MAP would increase from ~200 to ~500 mm/yr.
View in article
Zhang, C.F., Wang, Y., Li, Q., Wang, X.M., Deng, T., Tseng, Z.J., Takeuchi, G.T., Xie, G.P., Xu, Y.F. (2012) Diets and environments of late Cenozoic mammals in the Qaidam Basin, Tibetan Plateau: Evidence from stable isotopes. Earth and Planetary Science Letters 333, 70-82.
Show in contextTable S-2
View in article
Figures and Tables
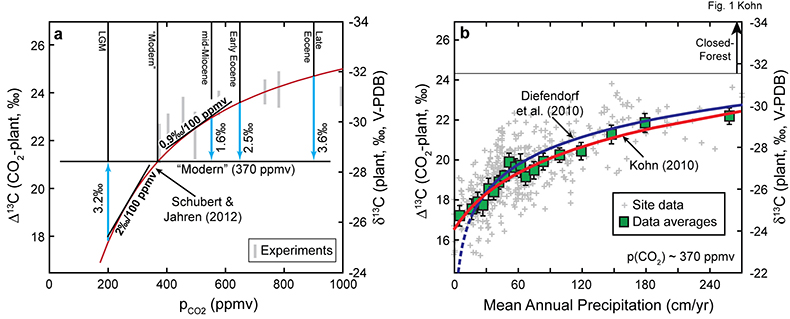
Figure 1 Proposed models for factors that influence δ13C of C3 plants. (a) pCO2. Differences are illustrated between geological conditions vs. AD 2000 (pCO2 = 370 ppmv, average δ13C = -28.5 for C3 biomass). LGM = Last Glacial Maximum. Note inverse relationship between δ13C and Δ13C. Experiments are for above-ground biomass (Schubert and Jahren, 2012), shifted to fit preferred curve. (b) Mean annual precipitation (data and data averages from Kohn, 2010).
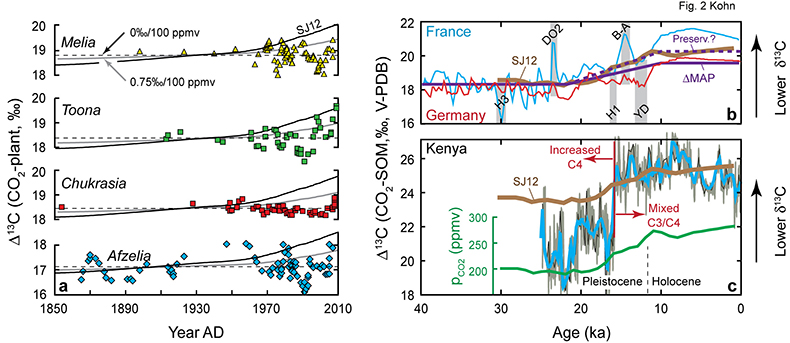
Figure 2 Tree ring and Pleistocene-Holocene records. (a) Tropical data from Thailand show near-zero pCO2 effect. Models are anchored on oldest 5-10 analyses (where any pCO2 effects should be smallest). SJ12 = model of Schubert and Jahren (2012). (b) Correction for changes in mean annual precipitation (“ΔMAP”), and possibly for organic matter preservation (“preserv.”) reproduces records independent of a direct pCO2-dependence. Brief climatic events also correlate with changes in Δ13C independent of pCO2. H1, H3 = Heinrich events 1 and 3; DO2 = Dansgaard-Oeschger event 2; B-A = Bølling-Allerød; YD = Younger Dryas. (c) An abrupt shift at c. 15.8 ka could reflect an abrupt threshold response of C4 biomass in the local ecosystem or variable contributions of diatoms to SOM (Fig. S-1).
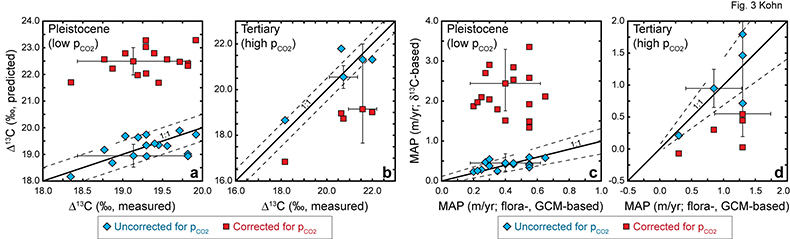
Figure 3 Cenozoic herbivore data. (a-d) Calculated vs. measured Δ13C, and calculated vs. independently estimated MAP, showing good correspondence with pCO2-independent model. Representative error bars (2σ) reflect propagated uncertainties in isotope compositions and tissue-diet fractionations, the dependence of δ13C on MAP, and proxy estimates of MAP and pCO2 (Table S-2).
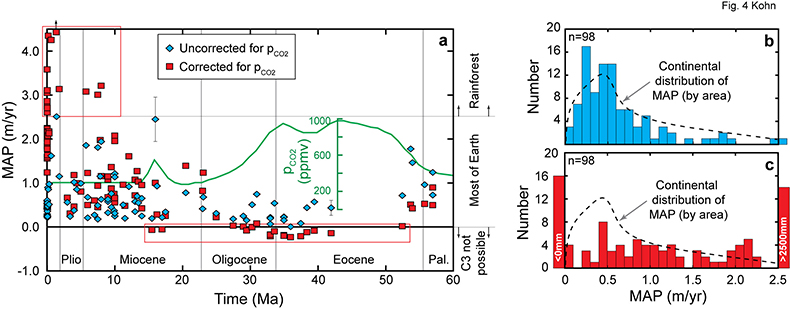
Figure 4 (a) Estimates of MAP for the Cenozoic based on fossil herbivore isotope compositions show a U-shaped distribution. pCO2-corrections imply widespread rainforests in the Pleistocene, and hyper-deserts in mid-Cenozoic. pCO2 curve simplified from Beerling and Royer (2011). (b-c) Histograms of Cenozoic MAP estimates show a better match between the global distribution (by area) of MAP (Kohn, 2010) vs. MAP estimated using the pCO2-independent model. The pCO2-dependent model predicts unusually high proportions of deserts (MAP < 0.1 m/yr) and rainforests (MAP > 2.5 m/yr) in the geologic record.
Back to article
Supplementary Figures and Tables
Table S-1 Specific regression results for leaf and tree ring δ13C.
| Leaf regressions | |||
| Parameter | Value ± 1σ | t | p |
| Intercept | 6.35 ± 2.07 | 3.06 | 0.002 |
| Elevation (m) | -1.80 ± 0.39x10-4 | -4.66 | <1x10-4 |
| log(MAP-300. mm) | 5.84 ± 0.24 | 24.85 | <1x10-4 |
| Abs(latitude. °) | 0.0139 ± 0.0042 | 3.28 | 0.001 |
| pCO2 (ppmv) | -0.0116 ± 0.0052 | -2.23 | 0.026 |
Δ13C = 6.35 - 1.80e-4 x Elev + 5.84*log(MAP-300) + 0.014 x Abs(lat.) – 0.012*pCO2
| Tree ring regressions* | Slope (±2σ) | pCO2 range (ppmv) |
| Thailand A. xylocarpa | 0.10 ± 0.68 | 285-390 |
| Thailand C. tabularis | 0.10 ± 0.54 | 285-385 |
| Thailand M. azedarach | -0.56 ± 1.07 | 300-390 |
| Thailand T. ciliata | 0.80 ± 1.11 | 300-390 |
| Cameroon B. cynometroides | 0.95 ± 0.69 | 300-385 |
| Cameroon B. eurycoma | 0.30 ± 0.83 | 295-375 |
| Cameroon D. ogea | 0.89 ± 0.67 | 280-390 |
| Cameroon T. ivorensis | 1.30 ± 0.53 | 285-385 |
| Bolivia C. ianeirensis | 1.25 ± 0.71 | 300-385 |
| Bolivia H. crepitans | 0.46 ± 1.16 | 290-385 |
| Bolivia S. fruticosa | 0.32 ± 0.70 | 290-390 |
| Average | 0.62 ± 1.05 | 285-390 |
| Borneo S. johorensis† | -0.04 ± 0.20 | 285-385 |
| Borneo S. superba† | 0.14 ± 0.29 | 285-370 |
| European composite† | -0.55 ± 0.67 | 300-370 |
*Regressions for specific tree ring data from van der Sleen et al. (2015) van der Sleen, P., Groenendijk, P., Vlam, M., Anten, N.P.R., Boom, A., Bongers, F., Pons, T.L., Terburg, G., Zuidema, P.A. (2015) No growth stimulation of tropical trees by 150 years of CO2 fertilization but water-use efficiency increased. Nature Geoscience 8, 24-28. (Thailand, Cameroon, Bolivia; multiple trees) and Loader et al. (2011 Loader, N.J., Walsh, R.P.D., Robertson, I., Bidin, K., Ong, R.C., Reynolds, G., McCarroll, D., Gagen, M., Young, G.H.F. (2011) Recent trends in the intrinsic water-use efficiency of ringless rainforest trees in Borneo. Philosophical Transactions of the Royal Society B-Biological Science 366, 3330-3339. ; Borneo; single trees). European composite is from Frank et al. (2015) Frank, D.C., Poulter, B., Saurer, M., Esper, J., Huntingford, C., Helle, G., Treydte, K., Zimmermann, N.E., Schleser, G.H., Ahlström, A., Ciais, P., Friedlingstein, P., Levis, S., Lomas, M., Sitch, S., Viovy, N., Andreu-Hayles, L., Bednarz, Z., Berninger, F., Boettger, T., D‘Alessandro, C.M., Daux, V., Filot, M., Grabner, M., Gutierrez, E., Haupt, M., Hilasvuori, E., Jungner, H., Kalela-Brundin, M., Krapiec, M., Leuenberger, M., Loader, N.J., Marah, H., Masson-Delmotte, V., Pazdur, A., Pawelczyk, S., Pierre, M., Planells, O., Pukiene, R., Reynolds-Henne, C.E., Rinne, K.T., Saracino, A., Sonninen, E., Stievenard, M., Switsur, V.R., Szczepanek, M., Szychowska-Krapiec, E., Todaro, L., Waterhouse, J.S., Weig, M. (2015) Water-use efficiency and transpiration across European forests during the Anthropocene. Nature Climate Change, doi: 10.1038/NCLIMATE2614. for Quercus and Pinus.
†Data not used in averages.
Table S-2 Herbivore isotope data and calculations of Δ13C and MAP.
| Source | Age (Ma) | δ13C (CO2. ‰) | pCO2 ppmv | δ13C (‰) | δ13C (plant. ‰) | Δ13C (K)* | Δ13C (S)* | Alt. (m) | Lat. (˚) | MAP (mm/yr) | Δ13C (K)* | Δ13C (S)* | MAP (K)* | MAP (S)* |
| Aiglstorfer et al. (2014) Aiglstorfer, M., Bocherens, H., Bohme, M. (2014) Large mammal ecology in the late Middle Miocene Gratkorn locality (Austria). Palaeobiodiversity and Palaeoenvironments 94, 189-213. | 12.1 | -6.1 | 300 | -12 | -26.25 | 20.69 | 21.79 | 350 | 47 | 919 | 1571 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 4 | -6.25 | 350 | -10 | -24.5 | 18.71 | 18.99 | 2000 | 35 | 377 | 456 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 6 | -6.2 | 300 | -9.75 | -23.75 | 17.98 | 19.07 | 2000 | 35 | 208 | 480 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 7 | -6.15 | 300 | -10 | -24 | 18.29 | 19.38 | 2000 | 35 | 274 | 581 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 7.5 | -6.1 | 300 | -9.5 | -23.5 | 17.82 | 18.91 | 2000 | 35 | 178 | 433 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 9 | -5.95 | 300 | -9.5 | -24 | 18.49 | 19.59 | 2000 | 35 | 322 | 655 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 9.5 | -6 | 300 | -10.5 | -24.5 | 18.96 | 20.06 | 2000 | 35 | 448 | 848 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 11.5 | -6 | 300 | -9.5 | -23.5 | 17.92 | 19.02 | 2000 | 35 | 197 | 463 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 13 | -5.85 | 300 | -10 | -24 | 18.6 | 19.69 | 2000 | 35 | 348 | 694 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 14 | -5.75 | 325 | -9.5 | -23.5 | 18.18 | 18.84 | 2000 | 35 | 250 | 414 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 17 | -5.4 | 550 | -9 | -23 | 18.01 | 16.19 | 2000 | 35 | 216 | -47 | |||
| Biasatti et al. (2010) Biasatti, D., Wang, Y., Deng, T. (2010) Strengthening of East Asian summer monsoon revealed by a shift in seasonal patterns in diet and climate after 2-3 Ma in northwest China. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 12-25. | 25 | -6 | 400 | -10 | -24 | 18.44 | 18.06 | 2000 | 35 | 310 | 225 | |||
| Biasatti et al. (2012) Biasatti, D., Wang, Y., Gao, F., Xu, Y., Flynn, L. (2012) Paleoecologies and paleoclimates of late Cenozoic mammals from Southwest China: evidence from stable carbon and oxygen isotopes. Journal of Asian Earth Science 44, 48-61. | 4 | -6.25 | 350 | -12 | -26 | 20.28 | 20.56 | 2000 | 25 | 1016 | 1169 | |||
| Biasatti et al. (2012) Biasatti, D., Wang, Y., Gao, F., Xu, Y., Flynn, L. (2012) Paleoecologies and paleoclimates of late Cenozoic mammals from Southwest China: evidence from stable carbon and oxygen isotopes. Journal of Asian Earth Science 44, 48-61. | 5 | -6.3 | 400 | -12 | -26 | 20.23 | 19.84 | 2000 | 25 | 990 | 809 | |||
| Biasatti et al. (2012) Biasatti, D., Wang, Y., Gao, F., Xu, Y., Flynn, L. (2012) Paleoecologies and paleoclimates of late Cenozoic mammals from Southwest China: evidence from stable carbon and oxygen isotopes. Journal of Asian Earth Science 44, 48-61. | 7.5 | -6.1 | 300 | -12.5 | -27 | 21.48 | 22.57 | 2000 | 25 | 1807 | 2934 | |||
| Biasatti et al. (2012) Biasatti, D., Wang, Y., Gao, F., Xu, Y., Flynn, L. (2012) Paleoecologies and paleoclimates of late Cenozoic mammals from Southwest China: evidence from stable carbon and oxygen isotopes. Journal of Asian Earth Science 44, 48-61. | 8 | -5.95 | 300 | -13 | -27 | 21.63 | 22.73 | 2000 | 25 | 1938 | 3136 | |||
| Bibi (2007) Bibi, F. (2007) Dietary niche partitioning among fossil bovids in late Miocene C3 habitats: Consilience of functional morphology and stable isotope analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 253, 529-538. | 8 | -5.95 | 300 | -10.75 | -25.25 | 19.8 | 20.89 | 450 | 33 | 630 | 1127 | |||
| Blondel et al. (1997) Blondel, C., Bocherens, H., Mariotti, A. (1997) Stable carbon and oxygen isotope ratios in ungulate teeth from French Eocene and Oligocene localities. Bulletin Societé Geologique de France 168, 775-781. | 29.5 | -6.3 | 540 | -9.8 | -24.05 | 18.19 | 16.44 | 50 | 44 | 153 | -71 | |||
| Blondel et al. (1997) Blondel, C., Bocherens, H., Mariotti, A. (1997) Stable carbon and oxygen isotope ratios in ungulate teeth from French Eocene and Oligocene localities. Bulletin Societé Geologique de France 168, 775-781. | 33 | -6 | 850 | -9 | -23.25 | 17.66 | 14.25 | 50 | 44 | 69 | -203 | |||
| Blondel et al. (1997) Blondel, C., Bocherens, H., Mariotti, A. (1997) Stable carbon and oxygen isotope ratios in ungulate teeth from French Eocene and Oligocene localities. Bulletin Societé Geologique de France 168, 775-781. | 35 | -6 | 950 | -9.9 | -24.15 | 18.6 | 14.85 | 50 | 44 | 233 | -178 | |||
| Blondel et al. (1997) Blondel, C., Bocherens, H., Mariotti, A. (1997) Stable carbon and oxygen isotope ratios in ungulate teeth from French Eocene and Oligocene localities. Bulletin Societé Geologique de France 168, 775-781. | 37.5 | -6.2 | 880 | -9.3 | -23.55 | 17.77 | 14.25 | 50 | 44 | 85 | -203 | |||
| Boardman and Secord (2013) Boardman, G.S., Secord, R. (2013) Stable isotope paleoecology of White River ungulates during the Eocene-Oligocene climate transition in northwestern Nebraska. Palaeogeography, Palaeoclimatology, Palaeoecology 375, 38-49. | 33 | -5.25 | 850 | -8.9 | -23.15 | 18.32 | 14.91 | 500 | 43 | 198 | -169 | |||
| Boardman and Secord (2013) Boardman, G.S., Secord, R. (2013) Stable isotope paleoecology of White River ungulates during the Eocene-Oligocene climate transition in northwestern Nebraska. Palaeogeography, Palaeoclimatology, Palaeoecology 375, 38-49. | 35 | -6 | 950 | -9.4 | -23.65 | 18.08 | 14.32 | 500 | 43 | 152 | -196 | |||
| Bocherens and Drucker (2003) Bocherens, H., Drucker, D. (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from Recent and ancient terrestrial ecosystems. International Journal of Osteoarcheology 13, 46-53. | 0.02 | -6.6 | 195 ± 10 | -20 | -25 | 18.87 | 22.42 | 50 | 44 | 250 ± 100 | 18.68 | 22.23 | 293 | 2075 |
| Bocherens et al. (2011) Bocherens, H., Drucker, D.G., Bonjean, D., Bridault, A., Conard, N.J., Cupillard, C., Germonpre, M., Honeisen, M., Munzel, S.C., Napierala, H., Patou-Mathis, M., Stephan, E., Uerpmann, H.P., Ziegler, R. (2011) Isotopic evidence for dietary ecology of cave lion (Panthera spelaea) in North-Western Europe: Prey choice, competition and implications for extinction. Quaternary International 245, 249-261. | 0.013 | -6.8 | 240 ± 10 | -20.5 | -25.5 | 19.19 | 21.53 | 1000 | 47 | 550 ± 200 | 19.64 | 21.98 | 412 | 1477 |
| Bocherens et al. (2011) Bocherens, H., Drucker, D.G., Bonjean, D., Bridault, A., Conard, N.J., Cupillard, C., Germonpre, M., Honeisen, M., Munzel, S.C., Napierala, H., Patou-Mathis, M., Stephan, E., Uerpmann, H.P., Ziegler, R. (2011) Isotopic evidence for dietary ecology of cave lion (Panthera spelaea) in North-Western Europe: Prey choice, competition and implications for extinction. Quaternary International 245, 249-261. | 0.013 | -6.8 | 240 ± 10 | -20.75 | -25.75 | 19.45 | 21.79 | 75 | 49 | 400 ± 200 | 19.36 | 21.69 | 426 | 1514 |
| Bocherens et al. (2011) Bocherens, H., Drucker, D.G., Bonjean, D., Bridault, A., Conard, N.J., Cupillard, C., Germonpre, M., Honeisen, M., Munzel, S.C., Napierala, H., Patou-Mathis, M., Stephan, E., Uerpmann, H.P., Ziegler, R. (2011) Isotopic evidence for dietary ecology of cave lion (Panthera spelaea) in North-Western Europe: Prey choice, competition and implications for extinction. Quaternary International 245, 249-261. | 0.034 | -6.7 | 210 ± 10 | -20.5 | -25.5 | 19.29 | 22.4 | 300 | 50 | 300 ± 100 | 18.93 | 22.04 | 391 | 2035 |
| Bocherens et al. (2011) Bocherens, H., Drucker, D.G., Bonjean, D., Bridault, A., Conard, N.J., Cupillard, C., Germonpre, M., Honeisen, M., Munzel, S.C., Napierala, H., Patou-Mathis, M., Stephan, E., Uerpmann, H.P., Ziegler, R. (2011) Isotopic evidence for dietary ecology of cave lion (Panthera spelaea) in North-Western Europe: Prey choice, competition and implications for extinction. Quaternary International 245, 249-261. | 0.034 | -6.7 | 210 ± 10 | -20.25 | -25.25 | 19.03 | 22.14 | 750 | 47 | 550 ± 200 | 19.69 | 22.8 | 356 | 1916 |
| Bocherens et al. (2015a) Bocherens, H., Drucker, D.G., Germonpre, M., Laznickova-Galetova, M., Naito, Y.I., Wissing, C., Bruzek, J., Oliva, M. (2015a) Reconstruction of the Gravettian food-web at Predmosti I using multi-isotopic tracking (C-13, N-15, S-34) of bone collagen. Quaternary International 359, 211-228. | 0.0305 | -6.7 | 205 ± 10 | -20.75 | -25.75 | 19.55 | 22.8 | 250 | 49 | 400 ± 200 | 19.32 | 22.57 | 466 | 2437 |
| Bocherens et al. (2015b) Bocherens, H., Hofman-Kaminska, E., Drucker, D.G., Schmolcke, U., Kowalczyk, R. (2015b) European Bison as a refugee species? Evidence from isotopic data on early Holocene Bison and other large herbivores in northern Europe. PLoS One 10, 10.1371/journal.pone.0115090. | 0.011 | -6.6 | 280 | -21 | -26 | 19.92 | 21.39 | 20 | 55 | 542 | 1199 | |||
| Britton et al. (2012) Britton, K., Gaudzinski-Windheuser, S., Roebroeks, W., Kindler, L., Richards, M.P. (2012) Stable isotope analysis of well-preserved 120,000-year-old herbivore bone collagen from the Middle Palaeolithic site of Neumark-Nord 2, Germany reveals niche separation between bovids and equids. Palaeogeography, Palaeoclimatology, Palaeoecology 333, 168-177. | 0.12 | -6.3 | 270 | -21.5 | -26.5 | 20.75 | 22.42 | 125 | 51 | 901 | 2011 | |||
| Castaños et al. (2014) Castaños, J., Zuluaga, M.C., Ortega, L.A., Murelaga, X., Alonso-Olazabal, A., Rofes, J., Castaños, P. (2014) Carbon and nitrogen stable isotopes of bone collagen of large herbivores from the Late Pleistocene Kiputz IX cave site (Gipuzkoa, north Iberian Peninsula) for palaeoenvironmental reconstruction. Quaternary International 339, 131-138. | 0.021 | -6.6 | 195 ± 10 | -21 | -26 | 19.92 | 23.46 | 200 | 43 | 550 ± 200 | 19.75 | 23.29 | 608 | 3337 |
| Cerling et al. (1997) Cerling, T.E., Harris, J.M., MacFadden, B.J., Leakey, M.G., Quade, J., Eisenmann, V., Ehleringer, J.R. (1997) Global vegetation change through the Miocene/Pliocene boundary. Nature 389, 153-158. | 14 | -5.75 | 325 | -10.5 | -24.5 | 19.22 | 19.89 | 1500 | 0 | 650 | 933 | |||
| Coltrain et al. (2004) Coltrain, J.B., Harris, J.M., Cerling, T.E., Ehleringer, J.R., Dearing, M.-D., Ward, J., Allen, J. (2004) Rancho La Brea stable isotope biogeochemistry and its implications for the palaeoecology of late Pleistocene, coastal Southern California. Palaeogeography, Palaeoclimatology, Palaeoecology 205, 199-219. | 0.012 | -6.7 | 265 | -21 | -26 | 19.82 | 21.59 | 50 | 34 | 602 | 1508 | |||
| Coltrain et al. (2004) Coltrain, J.B., Harris, J.M., Cerling, T.E., Ehleringer, J.R., Dearing, M.-D., Ward, J., Allen, J. (2004) Rancho La Brea stable isotope biogeochemistry and its implications for the palaeoecology of late Pleistocene, coastal Southern California. Palaeogeography, Palaeoclimatology, Palaeoecology 205, 199-219. | 0.0225 | -6.7 | 190 ± 10 | -20.5 | -25.5 | 19.29 | 22.99 | 50 | 34 | 450 ± 200 | 19.34 | 23.04 | 435 | 2827 |
| DeSantis et al. (2008) DeSantis, L.R.G., Wallace, S.C. (2008) Neogene forests from the Appalachians of Tennessee, USA: Geochemical evidence from fossil mammal teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 266, 59-68. | 5.75 | -6.2 | 300 | -13.5 | -27.5 | 21.9 | 23 | 900 | 36 | 1860 | 3015 | |||
| DeSantis et al. (2009) DeSantis, L.R.G., Feranec, R.S., MacFadden, B.J. (2009) Effects of Global Warming on Ancient Mammalian Communities and Their Environments. PLoS One 4, 10.1371/journal.pone.0005750. | 1.8 | -6.6 | 250 | -12.5 | -27 | 20.97 | 23.07 | 50 | 29 | 1152 | 3009 | |||
| Domingo et al. (2012) Domingo, L., Koch, P.L., Grimes, S.T., Morales J., Lopez-Martinez N. (2012) Isotopic paleoecology of mammals and the Middle Miocene Cooling event in the Madrid Basin (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 339, 98-113. | 14 | -5.75 | 325 | -9.75 | -23.75 | 18.44 | 19.11 | 1000 | 40 | 249 | 413 | |||
| Domingo et al. (2012) Domingo, L., Koch, P.L., Grimes, S.T., Morales J., Lopez-Martinez N. (2012) Isotopic paleoecology of mammals and the Middle Miocene Cooling event in the Madrid Basin (Spain). Palaeogeography, Palaeoclimatology, Palaeoecology 339, 98-113. | 15.5 | -5.25 | 550 ± 250 | -9 | -23 | 18.17 | 16.35 | 1000 | 40 | 300 ± 100 | 18.66 | 16.84 | 194 | -58 |
| Domingo et al. (2013) Domingo, L., Koch, P.L., Fernandez, M.H., Fox, D.L., Domingo, M.S., Alberdi, M.T. (2013) Late Neogene and early Quaternary paleoenvironmental and paleoclimatic conditions in southwestern Europe: Isotopic analyses on mammalian taxa. PLoS One 8, doi: 10.1371/journal.pone.0063739. | 9.5 | -6 | 300 | -10.5 | -24.75 | 19.23 | 20.32 | 1000 | 40 | 448 | 848 | |||
| Eberle et al. (2009) Eberle, J., Fricke, H., Humphrey, J. (2009) Lower-latitude mammals as year-round residents in Eocene Arctic forests. Geology 37, 499-502. | 52.5 | -5.9 | 715 ± 300 | -12 | -26 | 20.64 | 17.8 | 250 | 77 | 1300 ± 400 | 21.79 | 18.96 | 717 | 35 |
| Ecker et al. (2013) Ecker, M., Bocherens, H., Julien, M.A., Rivals, F., Raynal, J.P., Moncel, M.H. (2013) Middle Pleistocene ecology and Neanderthal subsistence: Insights from stable isotope analyses in Payre (Ardeche, southeastern France). Journal of Human Evolution 65, 363-373. | 0.175 | -7 | 200 | -12.5 | -27 | 20.55 | 23.95 | 200 | 45 | 856 | 4070 | |||
| Feranec and MacFadden (2006) Feranec, R.S., MacFadden, B.J. (2006) Isotopic discrimination of resource partitioning among ungulates in C3-dominated communities from the Miocene of Florida and California. Paleobiology 32, 191-205. | 9.5 | -6 | 300 | -11.7 | -25.7 | 20.22 | 21.31 | 50 | 33 | 762 | 1330 | |||
| Feranec et al. (2010) Feranec, R.S., Garcia, N., Diez, J.C., Arsuaga, J.L. (2010) Understanding the ecology of mammalian carnivorans and herbivores from Valdegoba cave (Burgos, northern Spain) through stable isotope analysis. Palaeogeography, Palaeoclimatology, Palaeoecology 297, 263-272. | 0.04 | -6.7 | 200 | -10.3 | -24.55 | 18.3 | 21.69 | 1000 | 42 | 215 | 1647 | |||
| Fox-Dobbs et al. (2008) Fox-Dobbs, K., Leonard, J.A., Koch, P.L. (2008) Pleistocene megafauna from eastern Beringia: paleoecological and paleoenvironmental interpretations of stable carbon and nitrogen isotope and radiocarbon records. Palaeogeography, Palaeoclimatology, Palaeoecology 261, 30-46. | 0.03 | -6.7 | 200 ± 10 | -21 | -26 | 19.82 | 23.21 | 300 | 65 | 275 ± 100 | 19.02 | 22.41 | 486 | 2672 |
| France et al. (2007) France, C.A.M., Zelanko, P.M., Kaufman, A.J., Holtz, T.R. (2007) Carbon and nitrogen isotopic analysis of Pleistocene mammals from the Saltville Quarry (Virginia, USA): Implications for trophic relationships. Palaeogeography, Palaeoclimatology, Palaeoecology 249, 271-282. | 0.016 | -6.8 | 230 ± 10 | -21 | -26 | 19.71 | 22.29 | 500 | 37 | 650 ± 200 | 19.9 | 22.48 | 584 | 2128 |
| Gilmour (2011) Gilmour, D.M. (2011) Chronology and ecology of late Pleistocene megafauna in the northern Willamette Valley, Oregon. MS (Portland State University, Portland, Oregon). | 0.014 | -6.8 | 240 | -21 | -26 | 19.71 | 22.05 | 50 | 45 | 520 | 1746 | |||
| Iacumin et al. (1997) Iacumin, P., Bocherens, H., Delgado Huertas, A., Mariotti, A., Longinelli, A. (1997) A stable isotope study of fossil mammal remains from the Paglicci Cave, southern Italy: N and C as palaeoenvironmental indicators. Earth and Planetary Science Letters 148, 349-357. | 0.023 | -6.7 | 195 ± 10 | -20.5 | -25.5 | 19.29 | 22.84 | 100 | 41 | 550 ± 200 | 19.74 | 23.29 | 412 | 2553 |
| Koch et al. (1995) Koch, P.L., Zachos, J.C., Dettman, D.L. (1995) Stable isotope stratigraphy and paleoclimatology of the Paleogene Bighorn Basin (Wyoming, USA). Palaeogeography, Palaeoclimatology, Palaeoecology 115, 61-89. | 54 | -5.5 | 600 ± 300 | -12.5 | -26.5 | 21.57 | 19.4 | 500 | 44 | 1300 ± 400 | 21.32 | 19.14 | 1467 | 454 |
| Koch et al. (1995) Koch, P.L., Zachos, J.C., Dettman, D.L. (1995) Stable isotope stratigraphy and paleoclimatology of the Paleogene Bighorn Basin (Wyoming, USA). Palaeogeography, Palaeoclimatology, Palaeoecology 115, 61-89. | 57 | -4.3 | 420 | -10 | -24 | 20.18 | 19.57 | 500 | 44 | 726 | 505 | |||
| Kocsis et al. (2014) Kocsis, L., Ozsvart, P., Becker, D., Ziegler, R., Scherler, L., Codrea, V. (2014) Orogeny forced terrestrial climate variation during the late Eocene-early Oligocene in Europe. Geology 42, 727-730. | 31 | -6 | 650 | -11.5 | -25.75 | 20.27 | 17.79 | 500 | 47 | 746 | 96 | |||
| Kocsis et al. (2014) Kocsis, L., Ozsvart, P., Becker, D., Ziegler, R., Scherler, L., Codrea, V. (2014) Orogeny forced terrestrial climate variation during the late Eocene-early Oligocene in Europe. Geology 42, 727-730. | 36 | -6 | 920 | -8.5 | -22.75 | 17.14 | 13.48 | 500 | 47 | 7 | -227 | |||
| Kohn and McKay (2012) Kohn, M.J., McKay, M.P. (2012) Paleoecology of late Pleistocene-Holocene faunas of eastern and central Wyoming, USA, with implications for LGM climate models. Palaeogeography, Palaeoclimatology, Palaeoecology 326-328, 42-53. | 0.02 | -6.6 | 195 ± 10 | -10 | -24.5 | 18.35 | 21.89 | 1500 | 45 | 200 ± 100 | 18.16 | 21.71 | 238 | 1855 |
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 23.1 | -5.9 | 300 | -11.55 | -25.55 | 20.16 | 21.25 | 100 | 45 | 681 | 1205 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 20.5 | -6.1 | 300 | -11.98 | -25.98 | 20.41 | 21.5 | 100 | 45 | 781 | 1359 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 27.5 | -6.3 | 450 | -10.22 | -24.22 | 18.36 | 17.42 | 100 | 45 | 185 | 36 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 30.3 | -6.1 | 650 | -11.27 | -25.27 | 19.66 | 17.18 | 100 | 45 | 507 | 5 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 33.6 | -5.25 | 875 | -10.54 | -24.54 | 19.78 | 16.27 | 100 | 45 | 543 | -86 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 38.4 | -6.2 | 850 | -11.15 | -25.15 | 19.44 | 16.02 | 100 | 45 | 439 | -106 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 39.5 | -6 | 850 | -11.36 | -25.36 | 19.86 | 16.45 | 100 | 45 | 572 | -71 | |||
| Kohn et al. (2015) Kohn, M.J., Strömberg, C.A.E., Madden, R.H., Dunn, R.E., Evans, S., Palacios, A., Carlini, A.A. (2015) Quasi-static Eocene-Oligocene climate in Patagonia promotes slow faunal evolution and mid-Cenozoic global cooling. Palaeogeography, Palaeoclimatology, Palaeoecology 435, 24-37. | 42 | -5.7 | 950 | -10.65 | -24.65 | 19.43 | 15.68 | 100 | 45 | 436 | -131 | |||
| Kuitems et al. (2015) Kuitems, M., van der Plicht, J., Druckerc, D.G., van Kolfschotena, T., Palstrab, S.W.L., Bocherens, H. (2015) Carbon and nitrogen stable isotopes of well-preserved Middle Pleistocene bone collagen from Schöningen (Germany) and their palaeoecological implications. Journal of Human Evolution, doi: 10.1016/j.jhevol.2015.1001.1008. | 0.285 | -7 | 225 | -21.25 | -26.25 | 19.77 | 22.48 | 150 | 52 | 515 | 2054 | |||
| Kuitems et al. (2015) Kuitems, M., van der Plicht, J., Druckerc, D.G., van Kolfschotena, T., Palstrab, S.W.L., Bocherens, H. (2015) Carbon and nitrogen stable isotopes of well-preserved Middle Pleistocene bone collagen from Schöningen (Germany) and their palaeoecological implications. Journal of Human Evolution, doi: 10.1016/j.jhevol.2015.1001.1008. | 0.3 | -6.9 | 275 | -22.25 | -27.25 | 20.92 | 22.49 | 150 | 52 | 979 | 2066 | |||
| Macfadden and Higgins (2004) MacFadden, B.J., Higgins, P. (2004) Ancient ecology of 15-million-year-old browsing mammals within C3 plant communities from Panama. Oecologia 140, 169-182. | 16 | -5.25 | 550 | -13 | -27 | 22.35 | 20.53 | 20 | 9 | 2459 | 1052 | |||
| Martin et al. (2011) Martin, C., Bentaleb, I., Antoine, P.O. (2011) Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 19-29. | 9 | -5.9 | 300 | -11 | -25 | 19.59 | 20.68 | 500 | 29 | 577 | 1046 | |||
| Martin et al. (2011) Martin, C., Bentaleb, I., Antoine, P.O. (2011) Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 19-29. | 13 | -5.9 | 300 | -11 | -25 | 19.59 | 20.68 | 500 | 29 | 577 | 1046 | |||
| Martin et al. (2011) Martin, C., Bentaleb, I., Antoine, P.O. (2011) Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 19-29. | 23 | -5.9 | 300 | -10.5 | -24.5 | 19.07 | 20.16 | 500 | 29 | 415 | 797 | |||
| Martin et al. (2011) Martin, C., Bentaleb, I., Antoine, P.O. (2011) Pakistan mammal tooth stable isotopes show paleoclimatic and paleoenvironmental changes since the early Oligocene. Palaeogeography, Palaeoclimatology, Palaeoecology 311, 19-29. | 29 | -6.3 | 500 | -10.25 | -24.25 | 18.4 | 16.98 | 500 | 29 | 249 | 16 | |||
| Maung-Thein et al. (2011) Maung-Thein, Z.-M. Takai, M., Uno, H., Wynn, J.G., Egi, N., Tsubamoto, T., Thaung-Htike, Aung-Naing-Soe, Maung-Maung, Nishimura, T., Yoneda, M. (2011) Stable isotope analysis of the tooth enamel of Chaingzauk mammalian fauna (late Neogene, Myanmar) and its implication to paleoenvironment and paleogeography. Palaeogeography, Palaeoclimatology, Palaeoecology 300, 11-22. | 6 | -6.2 | 300 | -11 | -25.5 | 19.81 | 20.9 | 300 | 22 | 676 | 1197 | |||
| Merceron et al. (2006) Merceron, G., Zazzo, A., Spassov, N., Geraads, D., Kovachev, D. (2006) Bovid paleoecology and paleoenvironments from the Late Miocene of Bulgaria: evidence from dental microwear and stable isotopes. Palaeogeography, Palaeoclimatology, Palaeoecology 241, 637-654. | 7.85 | -6 | 300 | -12 | -26.5 | 21.06 | 22.15 | 400 | 41 | 1155 | 1934 | |||
| Montanari et al. (2013) Montanari, S., Louys, J., Price, G.J. (2013) Pliocene paleoenvironments of southeastern Queensland, Australia inferred from stable isotopes of marsupial tooth enamel. PLoS One 8, doi: 10.1371/journal.pone.0066221. | 3.4 | -6.5 | 350 | -12.4 | -24.4 | 18.35 | 18.63 | 350 | 27 | 238 | 301 | |||
| Morgan et al. (1994) Morgan, M.E., Kingston, J.D., Marino, B.D. (1994) Carbon isotopic evidence for the emergence of C4 plants in the Neogene from Pakistan and Kenya. Nature 367, 162-165. | 10 | -6 | 300 | -12.5 | -26.5 | 21.06 | 22.15 | 450 | 34 | 1217 | 2029 | |||
| Nelson (2007) Nelson, S.V. (2007) Isotopic reconstructions of habitat change surrounding the extinction of Sivapithecus, a Miocene hominoid, in the Siwalik Group of Pakistan. Palaeogeography, Palaeoclimatology, Palaeoecology 243, 204-222. | 9 | -5.95 | 300 | -11.25 | -25.25 | 19.8 | 20.89 | 400 | 33 | 626 | 1121 | |||
| Passey et al. (2002) Passey, B.H., Cerling, T.E., Perkins, M.E., Voorhies, M.R., Harris, J.M., Tucker, S.T. (2002) Environmental change in the Great Plains; an isotopic record from fossil horses. Journal of Geology 110, 123-140. | 10 | -6 | 300 | -10 | -24 | 18.44 | 19.54 | 800 | 38 | 247 | 540 | |||
| Pushkina et al. (2014) Pushkina, D., Bocherens, H., Ziegler, R. (2014) Unexpected palaeoecological features of the Middle and Late Pleistocene large herbivores in southwestern Germany revealed by stable isotopic abundances in tooth enamel. Quaternary International 339, 164-178. | 0.035 | -6.7 | 200 ± 10 | -11.75 | -26 | 19.82 | 23.21 | 250 | 50 | 300 ± 100 | 18.93 | 22.33 | 547 | 2902 |
| Pushkina et al. (2014) Pushkina, D., Bocherens, H., Ziegler, R. (2014) Unexpected palaeoecological features of the Middle and Late Pleistocene large herbivores in southwestern Germany revealed by stable isotopic abundances in tooth enamel. Quaternary International 339, 164-178. | 0.6 | -6.7 | 250 | -13.5 | -28 | 21.91 | 24.02 | 250 | 50 | 1627 | 4092 | |||
| Qu et al. (2014) Qu, Y.T., Jin, C.Z., Zhang, Y.Q., Hu, Y.W., Shang, X., Wang, C.S. (2014) Preservation assessments and carbon and oxygen isotopes analysis of tooth enamel of Gigantopithecus blacki and contemporary animals from Sanhe Cave, Chongzuo, South China during the Early Pleistocene. Quaternary International 354, 52-58. | 1.4 | -6.6 | 250 | -15.5 | -28.5 | 22.54 | 24.65 | 200 | 22 | 2521 | 6131 | |||
| Raghavan et al. (2014) Raghavan, M., Themudo, G.E., Smith, C.I., Zazula, G., Campos, P.F. (2014) Musk ox (Ovibos moschatus) of the mammoth steppe: tracing palaeodietary and palaeoenvironmental changes over the last 50,000 years using carbon and nitrogen isotopic analysis. Quaternary Science Reviews 102, 192-201. | 0.021 | -6.6 | 195 ± 10 | -20.25 | -25.25 | 19.13 | 22.68 | 100 | 75 | 225 ± 100 | 18.95 | 22.5 | 264 | 1958 |
| Raghavan et al. (2014) Raghavan, M., Themudo, G.E., Smith, C.I., Zazula, G., Campos, P.F. (2014) Musk ox (Ovibos moschatus) of the mammoth steppe: tracing palaeodietary and palaeoenvironmental changes over the last 50,000 years using carbon and nitrogen isotopic analysis. Quaternary Science Reviews 102, 192-201. | 0.055 | -6.7 | 210 | -20.25 | -25.25 | 19.03 | 22.14 | 100 | 75 | 241 | 1529 | |||
| Richards and Hedges (2003) Richards, M.P., Hedges, R.E.M. (2003) Variations in bone collagen δ13C and d15N values of fauna from Northwest Europe over the last 40000 years. Palaeogeography, Palaeoclimatology, Palaeoecology 193, 261-267. | 0.035 | -6.7 | 200 ± 10 | -20 | -25 | 18.77 | 22.16 | 100 | 50 | 350 ± 100 | 19.18 | 22.57 | 254 | 1796 |
| Scherler et al. (2014) Scherler, L., Tütken, T., Becker, D. (2014) Carbon and oxygen stable isotope compositions of late Pleistocene mammal teeth from dolines of Ajoie (Northwestern Switzerland). Quaternary Research 82, 378-387. | 0.035 | -6.7 | 200 ± 10 | -11.6 | -25.6 | 19.4 | 22.79 | 600 | 48 | 450 ± 200 | 19.41 | 22.8 | 446 | 2521 |
| Scherler et al. (2014) Scherler, L., Tütken, T., Becker, D. (2014) Carbon and oxygen stable isotope compositions of late Pleistocene mammal teeth from dolines of Ajoie (Northwestern Switzerland). Quaternary Research 82, 378-387. | 0.08 | -6.5 | 220 | -11.5 | -25.5 | 19.5 | 22.34 | 500 | 48 | 470 | 2041 | |||
| Secord et al. (2008) Secord, R., Wing, S.L., Chew, A. (2008) Stable isotopes in early Eocene mammals as indicators of forest canopy structure and resource partitioning. Paleobiology 34, 282-300. | 53.75 | -5.6 | 620 ± 300 | -13 | -27 | 21.99 | 19.69 | 500 | 44 | 1300 ± 400 | 21.32 | 19.02 | 1784 | 546 |
| Secord et al. (2010) Secord, R., Wing, S.L., Chew, A. (2008) Stable isotopes in early Eocene mammals as indicators of forest canopy structure and resource partitioning. Paleobiology 34, 282-300. | 55.8 | -4.8 | 460 ± 150 | -11 | -25 | 20.72 | 19.68 | 500 | 44 | 1350 ± 400 | 21.4 | 20.35 | 965 | 541 |
| Secord et al. (2010) Secord, R., Wing, S.L., Chew, A. (2008) Stable isotopes in early Eocene mammals as indicators of forest canopy structure and resource partitioning. Paleobiology 34, 282-300. | 57 | -4.3 | 420 | -11 | -25 | 21.23 | 20.61 | 500 | 44 | 1246 | 913 | |||
| Segalen and Lee-Thorp (2009) Ségalen, L., Lee-Thorp, J.A. (2009) Palaeoecology of late Early Miocene fauna in the Namib based on 13C/12C and 18O/16O ratios of tooth enamel and ratite eggshell carbonate. Palaeogeography, Palaeoclimatology, Palaeoecology 277, 191-198. | 17.5 | -5.6 | 400 | -10 | -24 | 18.85 | 18.47 | 100 | 27 | 343 | 253 | |||
| Suraprasit et al. (2014) Suraprasit, K., Chaimanee, Y., Bocherens, H., Chavasseau, O., Jaeger, J.J. (2014) Systematics and phylogeny of middle Miocene Cervidae (Mammalia) from Mae Moh Basin (Thailand) and a paleoenvironmental estimate using enamel isotopy of sympatric herbivore species. Journal of Vertebrate Paleontology 34, 179-194. | 13.3 | -5.7 | 300 | -10.5 | -25 | 19.79 | 20.89 | 500 | 19 | 702 | 1238 | |||
| Tütken and Vennemann (2009) Tütken, T., Vennemann, T. (2009) Stable isotope ecology of Miocene large mammals from Sandelzhausen, southern Germany. Paläontologische Zeitschrift 83, 207-226. | 16.25 | -5.3 | 550 ± 250 | -11.5 | -25.5 | 20.73 | 18.91 | 450 | 49 | 850 ± 250 | 20.55 | 18.73 | 933 | 304 |
| Tütken et al. (2006) Tütken, T., Vennemann, T.W., Janz, H., Heizmann, E.P.J. (2006) Palaeoenvironment and palaeoclimate of the Middle Miocene lake in the Steinheim basin, SW Germany: a reconstruction from C, O, and Sr isotopes of fossil remains. Palaeogeography, Palaeoclimatology, Palaeoecology 241, 457-491. | 13.5 | -5.85 | 300 | -11.1 | -25.1 | 19.75 | 20.84 | 600 | 49 | 549 | 1003 | |||
| Tütken et al. (2013) Tütken, T., Kaiser, T.M., Vennemann, T., Merceron, G. (2013) Opportunistic feeding strategy for the earliest Old World hypsodont equids: Evidence from stable isotope and dental wear proxies. PLoS One 8, doi: 10.1371/journal.pone.0074463. | 10 | -6 | 300 ± 50 | -12.6 | -26.6 | 21.16 | 22.26 | 200 | 48 | 1200 ± 400 | 21.27 | 22.36 | 1141 | 1912 |
| Uno et al. (2011) Uno, K.T., Cerling, T.E., Harris, J.M., Kunimatsu, Y., Leakey, M.G., Nakatsukasa, M., Nakaya, H. (2011) Late Miocene to Pliocene carbon isotope record of differential diet change among East African herbivores. Proceedings of the National Academy of Sciences 108, 6509-6514. | 9.9 | -6 | 300 | -9.4 | -23.9 | 18.34 | 19.43 | 1500 | 1 | 369 | 726 | |||
| van Dam and Reichart (2009) van Dam, J.A., Reichart, G.J. (2009) Oxygen and carbon isotope signatures in late Neogene horse teeth from Spain and application as temperature and seasonality proxies. Palaeogeography, Palaeoclimatology, Palaeoecology 274, 64-81. | 3 | -6.4 | 350 | -11.5 | -25.5 | 19.6 | 19.88 | 900 | 40 | 559 | 659 | |||
| van Dam and Reichart (2009) van Dam, J.A., Reichart, G.J. (2009) Oxygen and carbon isotope signatures in late Neogene horse teeth from Spain and application as temperature and seasonality proxies. Palaeogeography, Palaeoclimatology, Palaeoecology 274, 64-81. | 7.5 | -6.1 | 300 | -11 | -25 | 19.38 | 20.48 | 900 | 40 | 490 | 912 | |||
| Wang and Deng (2005) Wang, Y., Deng, T. (2005) A 25 m.y. isotopic record of paleodiet and environmental change from fossil mammals and paleosols from the NE margin of the Tibetan Plateau. Earth and Planetary Science Letters 236, 322-338. | 10 | -6 | 300 | -10 | -24 | 18.44 | 19.54 | 2000 | 35 | 310 | 636 | |||
| White et al. (2009) White, T.D., Ambrose, S.H., Suwa, G., Su, D.F., DeGusta, D., Bernor, R.L., Boisserie, J.-R., Brunet, M., Delson, E., Frost, S., Garcia, N., Giaourtsakis, I.X., Haile-Selassie, Y., Howell, F.C., Lehmann, T., Likius, A., Pehlevan, C., Saegusa, H., Semprebon, G., Teaford, M., Vrba, E. (2009) Macrovertebrate paleontology and the Pliocene habitat of Ardipithecus ramidus. Science 326, 87-93. | 4.4 | -6.2 | 350 | -11.5 | -25.5 | 19.81 | 20.09 | 1600 | 9 | 849 | 984 | |||
| Zanazzi and Kohn (2008) Zanazzi, A., Kohn, M.J. (2008) Ecology and physiology of White River mammals based on stable isotope ratios of teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 257, 22-37. | 33 | -5.25 | 850 | -9.25 | -23.25 | 18.43 | 15.01 | 500 | 43 | 219 | -164 | |||
| Zanazzi and Kohn (2008) Zanazzi, A., Kohn, M.J. (2008) Ecology and physiology of White River mammals based on stable isotope ratios of teeth. Palaeogeography, Palaeoclimatology, Palaeoecology 257, 22-37. | 35 | -6 | 950 | -10 | -24 | 18.44 | 14.69 | 500 | 43 | 221 | -180 | |||
| Zhang et al. (2012) Zhang, C.F., Wang, Y., Li, Q., Wang, X.M., Deng, T., Tseng, Z.J., Takeuchi, G.T., Xie, G.P., Xu, Y.F. (2012) Diets and environments of late Cenozoic mammals in the Qaidam Basin, Tibetan Plateau: Evidence from stable isotopes. Earth and Planetary Science Letters 333, 70-82. | 11.25 | -6 | 300 | -10.5 | -24.5 | 18.96 | 20.06 | 3000 | 37 | 500 | 928 |
*K = model of Kohn (2010) Kohn, M.J. (2010) Carbon isotope compositions of terrestrial C3 plants as indicators of (paleo)ecology and (paleo)climate. Proceedings of the National Academy of Sciences 107, 19691-19695. (pCO2-independent); S = model of Schubert and Jahren (2012) Schubert, B.A., Jahren, A.H. (2012) The effect of atmospheric CO2 concentration on carbon isotope fractionation in C3 land plants. Geochimica et Cosmochimica Acta 96, 29-43. (pCO2-dependent)
Note: Uncertainties in δ13C of atmospheric CO2 for the pre-Pleistocene is ~±0.35 ‰ (Tipple et al., 2010 Tipple, B.J., Meyers, S.R., Pagani, M. (2010) Carbon isotope ratio of Cenozoic CO2: a comparative evaluation of available geochemical proxies. Paleoceanography 25, doi: 10.1029/2009PA001851. ).
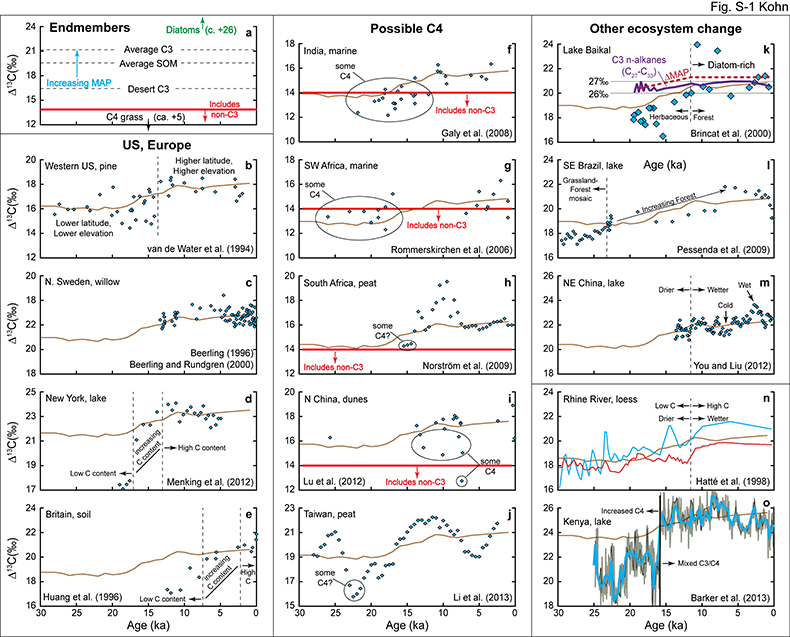
Figure S-1 (a) Principles of carbon isotope compositions relevant to interpreting isotopic records of sediment organic matter (SOM). C4 grasses have extremely low Δ13C, C3 plants have increasing Δ13C with increasing MAP, and diatoms have extremely high Δ13C. Applying a ~2 ‰ offset between plants and SOM (Bowling et al., 2008) implies a minimum cutoff of Δ13C(CO2-SOM) ~ 14 ‰ for which other sources of carbon besides C3 plants must be considered. (b-o) Records of Pleistocene-Holocene transition annotated with observations and interpretations from the original publications.
Table S-3 Calculated pCO2-dependencies of δ13C in C3 plants.
| Record | pCO2 coefficient ±2 s.e. | pCO2 range |
| (‰/100 ppmv) | (ppmv, to nearest 5 ppmv) | |
| Modern leaves | -1.2 ± 1.0 | 325-380 |
| Tropical tree rings | 0.62 ± 1.05 | 285-390 |
| Borneo trees† | (0.0 ± 0.2) | 285-385 |
| European tree rings† | (-0.55 ± 0.67) | 300-370 |
| Sediment records† | (≤1.0 or ~0.0) | 185-280 |
| Pleistocene fossils* | -0.03 ± 0.11 | 190 to 370 |
| Paleogene fossils* | -0.03 ± 0.24 | 370 to 715 |
| Weighted average | -0.04 ± 0.26 | 190-715 |
†Not used in weighted average.
*Calculations are referenced to conditions at 2000 AD, which had pCO2 = 370 ppmv
Table S-4 Summary of records and factors influencing calculations.
| Data source | Largest uncertainties affecting calculations (magnitude, ‰) |
| Modern leaves | Data scatter (1-2 ‰) |
| Experiments | Poorly constrained MAP, plastic phenotypic response (unknown) |
| Tropical tree rings | Oscillations of unknown origin (1-2 ‰) |
| Pleistocene-Holocene sediments | MAP, C3/C4 changes, variable diatoms, variable C preservation, oscillations of unknown origin (2-5 ‰) |
| Quaternary fossils | Data scatter, uncertain tissue-diet fractionation (<1 ‰) |
| Tertiary fossils | Poorly constrained pCO2, MAP (<1 ‰) |