Lanthanum anomalies provide constraints on macrofaunal predation at methane seeps
Affiliations | Corresponding Author | Cite as | Funding informationPublished by the European Association of Geochemistry
under Creative Commons License CC BY-NC-ND 4.0
Keywords: submarine seep ecosystems, methanotrophy-dependent mussels, predation, lanthanum

- Share this article




-
Article views:649Cumulative count of HTML views and PDF downloads.
- Download Citation
- Rights & Permissions

top
Abstract

Figures
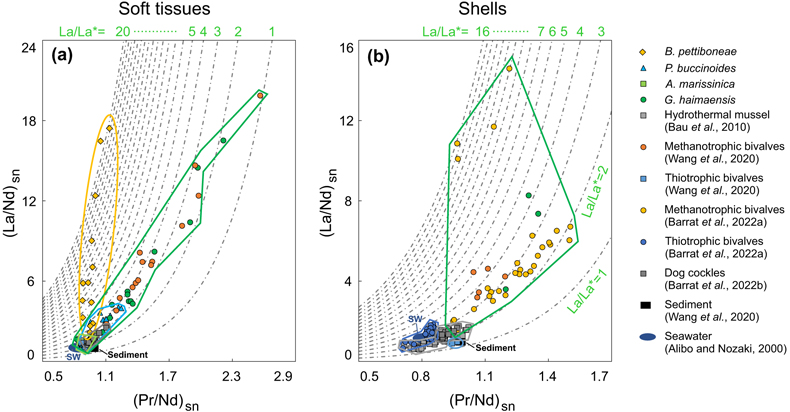
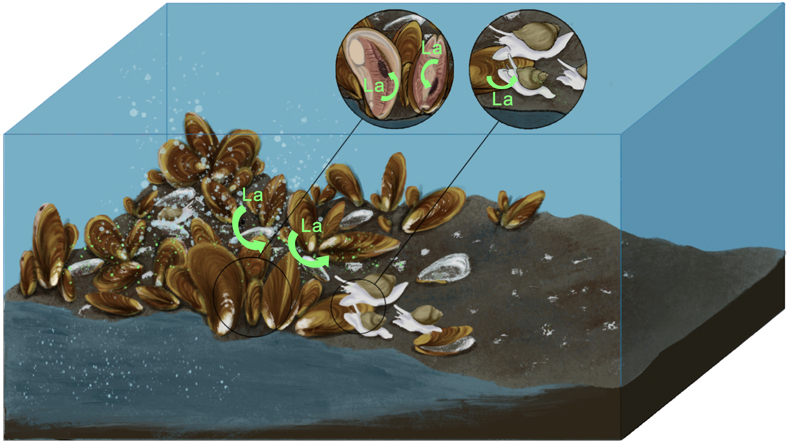
 Figure 1 (a) Approximate sampling location (Haima seeps). (b) Coexisting Gigantidas haimaensis (periphery) and Phymorhynchus buccinoides (centre) on the seabed. (c) Species investigated in this study. (c1-c2) Archivesica marissinica; (c3-c4) G. haimaensis; (c5) Branchipolynoe pettiboneae; (c6) P. buccinoides. Note that in c4, B. pettiboneae is parasitic in G. haimaensis. Scale bars are 3 cm. | 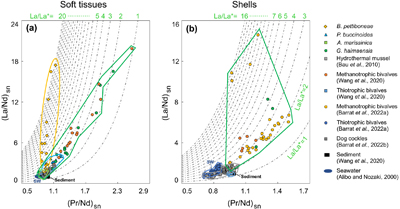 Figure 2 (La/Nd)sn vs. (Pr/Nd)sn plots for (a) soft tissues and (b) shells of various macrofauna from the Haima seeps. Previous work (Bau et al., 2010; Wang et al., 2020; Barrat et al., 2022a,b), seawater (SW; Alibo and Nozaki, 2000) and sediment from the Haima seeps (Wang et al., 2020) are shown for comparison. | 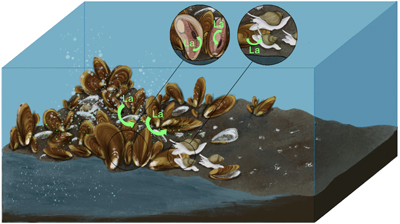 Figure 3 Conceptual diagram of predator-prey interactions within seep ecosystems. La = lanthanum. |
| Figure 1 | Figure 2 | Figure 3 |
top
Introduction
Understanding food web structures is essential for comprehending the composition, functionality, and stability of ecosystems. Predation is a crucial component of food web structures and plays a vital role in regulating the population dynamics and maintaining population equilibrium within an ecosystem. Predator–prey interactions also influence species distribution across trophic levels, affecting the flow of energy and matter within the ecosystem. These interactions, in conjunction with adaptation and evolution, contribute to the complexity of food webs and ecological networks. Overall, predator-prey relationships are recognised as important mechanisms in structuring ecosystems (van Denderen et al., 2018
van Denderen, P.D., Lindegren, M., MacKenzie, B.R., Watson, R.A., Andersen, K.H. (2018) Global patterns in marine predatory fish. Nature Ecology & Evolution 2, 65–70. https://doi.org/10.1038/s41559-017-0388-z
).Submarine seep ecosystems are found worldwide along continental margins (Levin et al., 2016
Levin, L.A., Baco, A.R., Bowden, D.A., Colaco, A., Cordes, E.E., Cunha, M.R., Demopoulos, A.W.J., Gobin, J., Grupe, B.M., Le, J., Metaxas, A., Netburn, A.N., Rouse, G.W., Thurber, A.R., Tunnicliffe, V., Van Dover, C.L., Vanreusel, A., Watling, L. (2016) Hydrothermal vents and methane seeps: rethinking the sphere of influence. Frontiers in Marine Science 3, 72. https://doi.org/10.3389/fmars.2016.00072
). Unlike ecosystems that rely on photosynthesis, seeps are typified by chemosynthesis-based ecosystems that primarily obtain energy from the oxidation of reduced compounds like methane and hydrogen sulfide. These ecosystems are typified by a low diversity but high abundance of the dominant species (Jørgensen and Boetius, 2007Jørgensen, B.B., Boetius, A. (2007) Feast and famine — microbial life in the deep-sea bed. Nature Reviews Microbiology 5, 770–781. https://doi.org/10.1038/nrmicro1745
). The community structure is influenced by the availability of energy sources, as well as other factors such as predation, water depth, substrate type, and ecological succession (MacAvoy et al., 2002MacAvoy, S.E., Carney, R.S., Fisher, C.R., Macko, S.A. (2002) Use of chemosynthetic biomass by large, mobile, benthic predators in the Gulf of Mexico. Marine Ecology Progress Series 225, 65–78. https://doi.org/10.3354/meps225065
; Morganti et al., 2022Morganti, T.M., Slaby, B.M., de Kluijver, A., Busch, K., Hentschel, U., Middleburg, J.J., Grotheer, H., Mollenhauer, G., Dannheim, J., Rapp, H.T., Purser, A., Boetius, A. (2022) Giant sponge grounds of Central Arctic seamounts are associated with extinct seep life. Nature Communications 13, 638. https://doi.org/10.1038/s41467-022-28129-7
).The impact of predation on the community structure of seep ecosystems is still not fully understood (Levin et al., 2016
Levin, L.A., Baco, A.R., Bowden, D.A., Colaco, A., Cordes, E.E., Cunha, M.R., Demopoulos, A.W.J., Gobin, J., Grupe, B.M., Le, J., Metaxas, A., Netburn, A.N., Rouse, G.W., Thurber, A.R., Tunnicliffe, V., Van Dover, C.L., Vanreusel, A., Watling, L. (2016) Hydrothermal vents and methane seeps: rethinking the sphere of influence. Frontiers in Marine Science 3, 72. https://doi.org/10.3389/fmars.2016.00072
). Previous studies have suggested that predators in these ecosystems do not play a dominate role and their influence is typically considered modest (Portail et al., 2016Portail, M., Olu, K., Dubois, S.F., Escobar-Briones, E., Gelinas, Y., Menot, L., Sarrazin, J. (2016) Food-web complexity in Guaymas Basin hydrothermal vents and cold seeps. PLoS One 11, e0162263. https://doi.org/10.1371/journal.pone.0162263
). However, some studies have shown that predation can facilitate trophic transfer and regulate species diversity in seep ecosystems (MacAvoy et al., 2002MacAvoy, S.E., Carney, R.S., Fisher, C.R., Macko, S.A. (2002) Use of chemosynthetic biomass by large, mobile, benthic predators in the Gulf of Mexico. Marine Ecology Progress Series 225, 65–78. https://doi.org/10.3354/meps225065
; Olsen et al., 2014Olsen, B.R., Troedsson, C., Hadziavdic, K., Pederson, R.B., Rapp, H.T. (2014) A molecular gut content study of Themisto abyssorum (Amphipoda) from Arctic hydrothermal vent and cold seep systems. Molecular Ecology 23, 3877–3889. https://doi.org/10.1111/mec.12511
). For example, predation pressure has been found to be the main factor behind the inverse correlation between macrofauna and meiofauna/nematode densities among the seep benthos (Van Gaever et al., 2009Van Gaever, S., Galeron, J., Sibuet, M., Vanreusel, A. (2009) Deep-sea habitat heterogeneity influence on meiofaunal communities in the Gulf of Guinea. Deep-Sea Research II 56, 2259–2269. https://doi.org/10.1016/j.dsr2.2009.04.008
). Predation by metazoans may also explain the differences in copepod abundance between mussel beds and tubeworm colonies at seeps in the Gulf of Mexico (Plum et al., 2015Plum, C., Gollner, S., Martínez-Arbizu, P., Bright, M. (2015) Diversity and composition of the copepod communities associated with megafauna around a cold seep in the Gulf of Mexico with remarks on species biogeography. Marine Biodiversity 45, 419–432. https://doi.org/10.1007/s12526-014-0310-8
). Moreover, seeps serve as nurseries for deep sea predators, which can greatly affect the diversity of animal species along continental margins (Treude et al., 2011Treude, T., Kiel, S., Linke, P., Peckmann, J., Goedert, J.L. (2011) Elasmobranch egg capsules associated with modern and ancient cold seeps: a nursery for marine deep-water predators. Marine Ecology Progress Series 437, 175–181. https://doi.org/10.3354/meps09305
; Danovaro et al., 2022Danovaro, R., Fanelli, E., Aguzzi, J., Billett, D., Carugati, L., Corinaldesi, C., Dell’Anno, A., Ggerde, K., Jamieson, A.J., Kark, S., McClain, C., Levin, L., Levin, N., Ramirez-Llodra, E., Ruhl, H., Smith, C.R., Snelgrove, P.V.R., Thomsen, L., Van Dover, C.L., Yasuhara, M. (2022) Ecological variables for developing a global deep-ocean monitoring and conservation strategy. Nature Ecology & Evolution 4, 181–192. https://doi.org/10.1038/s41559-019-1091-z
). Understanding the variables influencing the community composition of seep ecosystems, particularly the role of predation, is crucial for comprehending the succession dynamics and the evolution of chemosynthesis-based faunas in Earth history (Treude et al., 2011Treude, T., Kiel, S., Linke, P., Peckmann, J., Goedert, J.L. (2011) Elasmobranch egg capsules associated with modern and ancient cold seeps: a nursery for marine deep-water predators. Marine Ecology Progress Series 437, 175–181. https://doi.org/10.3354/meps09305
; Kiel et al., 2016Kiel, S., Amano, K., Jenkins, R.G. (2016) Predation scar frequencies in chemosymbiotic bivalves at an Oligocene seep deposit and their potential relation to inferred sulfide tolerances. Palaeogeography, Palaeoclimatology, Palaeoecology 453, 139–145. https://doi.org/10.1016/j.palaeo.2016.04.026
).Bathymodiolin mussels serve as an ideal focal point for investigating the trophic transfer and the turnover of material within seep ecosystems. Regarding their trophic level, mussels can be classified as primary consumers (Portail et al., 2016
Portail, M., Olu, K., Dubois, S.F., Escobar-Briones, E., Gelinas, Y., Menot, L., Sarrazin, J. (2016) Food-web complexity in Guaymas Basin hydrothermal vents and cold seeps. PLoS One 11, e0162263. https://doi.org/10.1371/journal.pone.0162263
). Seep dwelling mussels rely on endosymbiotic primary producers, such as methanotrophic and/or thiotrophic bacteria. In turn, mussels serve as prey for higher level consumers. Representative examples of these higher level consumers include the turrid gastropod Phymorhynchus buccinoides and the scale worm Branchipolynoe pettiboneae at the Haima seeps of the South China Sea (Fujikura et al., 2009Fujikura, K., Sasaki, T., Yamanaka, T., Yoshida, T. (2009) Turrids whelk, Phymorhynchus buccinoides feeds on Bathymodiolus mussels at a seep site in Sagami Bay, Japan. Plankton and Benthos Research 4, 23–30. https://doi.org/10.3800/pbr.4.23
; Takahashi et al., 2012Takahashi, Y., Sasaki, Y., Chikaraishi, Y., Tsuchiya, M., Watanabe, H., Asahida, T., Maruyama, T., Fujikura, K. (2012) Does the symbiotic scale-worm feed on the host mussel in deep-sea vent fields? Researches in Organic Geochemistry 28, 23–26.
).Turrid gastropods of the genus Phymorhynchus are common in various deep sea habitats such as hydrothermal vents, hydrocarbon seeps, and sunken wood, where they are unspecialised predators that benefit from the high mass of available food in these ecosystems (Sasaki et al., 2010
Sasaki, T., Warén, A., Kano, Y., Okutani, T., Fujikura, K. (2010) Gastropods from recent hot vents and cold seeps: systematics, diversity and life strategies. In: Kiel, S. (Ed.) The Vent and Seep Biota. Springer, Dordrecht, 169–254. https://doi.org/10.1007/978-90-481-9572-5_7
). Stable carbon and nitrogen isotope analysis conducted on P. buccinoides specimens collected at the Haima seeps indicated a dependence on local chemosynthetic carbon sources, with the gastropods taking up methane-derived carbon through predation (Ke et al., 2022Ke, Z., Li, R., Chen, Y., Chen, D., Chen, Z., Lian, X., Tan, Y. (2022) A preliminary study of macrofaunal communities and their carbon and nitrogen stable isotopes in the Haima cold seeps, South China Sea. Deep-Sea Research I 184, 103774. https://doi.org/10.1016/j.dsr.2022.103774
). Phymorhynchus has a well developed, funnel-shaped rhynchostome that facilitates its predatory behaviour (Warén and Bouchet, 2001Warén, A., Bouchet, P. (2001) Gastropoda and Monoplacophora from hydrothermal vents and seeps; new taxa and records. The Veliger 44, 116–231.
), and fragments of various organisms in the gut of Phymorhynchus have been documented (Sasaki et al., 2010Sasaki, T., Warén, A., Kano, Y., Okutani, T., Fujikura, K. (2010) Gastropods from recent hot vents and cold seeps: systematics, diversity and life strategies. In: Kiel, S. (Ed.) The Vent and Seep Biota. Springer, Dordrecht, 169–254. https://doi.org/10.1007/978-90-481-9572-5_7
). Its predation on mussels has been demonstrated by in situ behavioural observations and bait trap experiments as well as anatomic examination (Fujikura et al., 2009Fujikura, K., Sasaki, T., Yamanaka, T., Yoshida, T. (2009) Turrids whelk, Phymorhynchus buccinoides feeds on Bathymodiolus mussels at a seep site in Sagami Bay, Japan. Plankton and Benthos Research 4, 23–30. https://doi.org/10.3800/pbr.4.23
).Scale worms can thrive in a wide range of habitats and frequently engage in a ‘parasitic’ relationship with mussels within seep ecosystems (Becker et al., 2013
Becker, E.L., Cordes, E.E., Macko, S.A., Lee, R.W., Fisher, C.R. (2013) Using stable isotope compositions of animal tissues to infer trophic interactions in Gulf of Mexico lower slope seep communities. PLoS One 8, e74459. https://doi.org/10.1371/journal.pone.0074459
; Yao et al., 2022Yao, G., Zhang, H., Xiong, P., Jia, H., He, M. (2022) Effects of scale worm parasitism on interactions between the symbiotic gill microbiome and gene regulation in deep sea mussel hosts. Frontiers in Microbiology 13, 940766. https://doi.org/10.3389/fmicb.2022.940766
). Previous stable isotope analyses conducted on scale worms from hydrocarbon seeps indicated their reliance on chemosynthesis-based biomass (Becker et al., 2013Becker, E.L., Cordes, E.E., Macko, S.A., Lee, R.W., Fisher, C.R. (2013) Using stable isotope compositions of animal tissues to infer trophic interactions in Gulf of Mexico lower slope seep communities. PLoS One 8, e74459. https://doi.org/10.1371/journal.pone.0074459
; Ke et al., 2022Ke, Z., Li, R., Chen, Y., Chen, D., Chen, Z., Lian, X., Tan, Y. (2022) A preliminary study of macrofaunal communities and their carbon and nitrogen stable isotopes in the Haima cold seeps, South China Sea. Deep-Sea Research I 184, 103774. https://doi.org/10.1016/j.dsr.2022.103774
). Based on their jaw structure, it has been inferred that the scale worms exhibit predatory behaviour (Fauchald and Jumars, 1979Fauchald, K., Jumars, P.A. (1979) The diet of worms: a study of polychaete feeding guilds. Oceanography and Marine Biology Annual Review 17, 193–284.
). Likewise, mussel fragments have been discovered among the stomach contents of the parasitic scale worm Branchipolynoe symmyitilida (Desbruyères et al., 1985Desbruyères, D., Gaill, F., Laubier, L., Fouquet, Y. (1985) Polychaetous annelids from hydrothermal vent ecosystems: an ecological overview. Bulletin of the Biological Society of Washington 6, 103–116.
). Nitrogen stable isotope compositions of amino acids of the scale worm B. pettiboneae and its host mussel indicate that the mussel serves as the primary source of amino acids for the adult parasitic worm (Takahashi et al., 2012Takahashi, Y., Sasaki, Y., Chikaraishi, Y., Tsuchiya, M., Watanabe, H., Asahida, T., Maruyama, T., Fujikura, K. (2012) Does the symbiotic scale-worm feed on the host mussel in deep-sea vent fields? Researches in Organic Geochemistry 28, 23–26.
). Parasitism of B. pettiboneae within G. haimaensis was confirmed and described in detail by Yao et al. (2022)Yao, G., Zhang, H., Xiong, P., Jia, H., He, M. (2022) Effects of scale worm parasitism on interactions between the symbiotic gill microbiome and gene regulation in deep sea mussel hosts. Frontiers in Microbiology 13, 940766. https://doi.org/10.3389/fmicb.2022.940766
.Here, we developed a new approach to quantify the trophic transfer among seep macrofauna (the thiotrophy-dependent vesicomyid clam Archivesica marissinica, the aerobic methanotrophy-dependent bathymodiolin mussel Gigantidas haimaensis, the heterotrophic turrid gastropod P. buccinoides, and the scale worm B. pettiboneae) from the Haima seeps of the South China Sea (Fig. 1, Table S-1), with the latter two species known to be feeding on mussels. We analysed the contents of rare earth elements (REEs), particularly lanthanum (La), of these species to evaluate the predation dynamics in seep ecosystems. This approach is based on the facts that (1) the occurrence of La anomalies in seep dwelling metazoans is a robust fingerprint of methanotrophy since the second step in the aerobic oxidation of methane (CH3OH → HCHO) can be catalysed by La (Semrau et al., 2018
Semrau, J.D., Dispirito, A.A., Gu, W., Yoon, S. (2018) Metals and methanotrophy. Applied and Environmental Microbiology 84, e02289–17. https://doi.org/10.1128/AEM.02289-17
; Wang et al., 2020Wang, X., Barrat, J.-A., Bayon, G., Chauvaud, L., Feng, D. (2020) Lanthanum anomalies as fingerprints of methanotrophy. Geochemical Perspectives Letters 14, 26–30. https://doi.org/10.7185/geochemlet.2019
), and (2) aerobic methanotrophy-dependent mussels are ubiquitous in seep ecosystems (Wang et al., 2022Wang, X., Guan, H., Qiu, J.-W., Xu, T., Peckmann, J., Chen, D., Feng, D. (2022) Macro-ecology of cold seeps in the South China Sea. Geosystems and Geoenvironment 1, 100081. https://doi.org/10.1016/j.geogeo.2022.100081
).Figure 1 (a) Approximate sampling location (Haima seeps). (b) Coexisting Gigantidas haimaensis (periphery) and Phymorhynchus buccinoides (centre) on the seabed. (c) Species investigated in this study. (c1-c2) Archivesica marissinica; (c3-c4) G. haimaensis; (c5) Branchipolynoe pettiboneae; (c6) P. buccinoides. Note that in c4, B. pettiboneae is parasitic in G. haimaensis. Scale bars are 3 cm.
top
Results
The contents of light REEs in the four examined metazoan species vary from several ng/g to several hundred ng/g. Notably, only two samples of bathymodiolin mussels exhibit contents at the μg/g level (Tables S-2 to S-5). The distribution patterns of REEs in the chemosymbiotic bivalves align with previous studies – the REE compositions of the clams fall between the compositions of terrigenous sediments and seawater, lacking light REE enrichment (Fig. S-2), while the light REE contents in the gills of the mussels are one order of magnitude higher than contents in the clams, particularly with respect to La (Fig. S-3). The scale worms as a whole and some of types of soft tissue (gills and visceral mass) of the turrid gastropods reveal an enrichment of La (Figs. S-4, S-5). However, no such enrichment was observed in the shells of the turrid gastropods (Fig. S-4).
top
Discussion
By determining the La and Ce anomalies (La/La* and Ce/Ce*; Eq. 1 and 2; cf. Barrat et al., 2023
Barrat, J.-A., Bayon, G., Lalonde, T. (2023) Calculation of cerium and lanthanum anomalies in geological and environmental samples. Chemical Geology 615, 121202. https://doi.org/10.1016/j.chemgeo.2022.121202
)Eq. 1
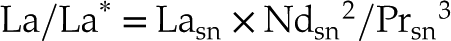
Eq. 2
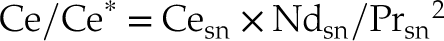
for four invertebrate species from the Haima seeps, we find that the REE patterns of the thiotrophy-dependent A. marissinica resemble the REE compositions of other seep dwelling clams (Wang et al., 2020
Wang, X., Barrat, J.-A., Bayon, G., Chauvaud, L., Feng, D. (2020) Lanthanum anomalies as fingerprints of methanotrophy. Geochemical Perspectives Letters 14, 26–30. https://doi.org/10.7185/geochemlet.2019
). Additionally, the La anomaly (La/La* = 1.15–2.64) observed for A. marissinica is similar to that of thiotrophy-dependent mussels from seeps on the Costa Rica margin (Barrat et al., 2022aBarrat, J.-A., Bayon, G., Carney, R.S., Chauvaud, L. (2022a) Rare earth elements as new biogeochemical proxies in deep-sea mussels. Chemical Geology 610, 121102. https://doi.org/10.1016/j.chemgeo.2022.121102
). The Ce anomaly in A. marissinica soft tissue (Ce/Ce* = 0.23–1.02) is typically lower than that of its shells (Ce/Ce* = 1.08–1.42). The presence of positive Ce anomalies in shells, agreeing with reducing conditions, is likely indicative of the clams’ semi-infaunal lifestyle. In contrast, the shells of the epifaunal mussel G. haimaensis yielded minor negative Ce anomalies (Ce/Ce* = 0.90–1.03), reflecting their exposure to oxic seawater. The La anomalies of mussel shells (La/La* = 2.10–3.71) are similar to previous findings (La/La* = 2.50–3.92, Wang et al., 2020Wang, X., Barrat, J.-A., Bayon, G., Chauvaud, L., Feng, D. (2020) Lanthanum anomalies as fingerprints of methanotrophy. Geochemical Perspectives Letters 14, 26–30. https://doi.org/10.7185/geochemlet.2019
) – slightly higher than the La anomalies of Gigantidas shells from the Brine Pool and Bush Hill sites of the Gulf of Mexico (La/La* = 1.73–2.70), but significantly lower than the anomalies of two Bathymodiolus shells from Edison Seamount (La/La* = 7.88–11.89; Barrat et al., 2022aBarrat, J.-A., Bayon, G., Carney, R.S., Chauvaud, L. (2022a) Rare earth elements as new biogeochemical proxies in deep-sea mussels. Chemical Geology 610, 121102. https://doi.org/10.1016/j.chemgeo.2022.121102
). While the magnitude of the mussels’ La anomaly could possibly relate to the efficiency of La utilisation during aerobic oxidation of methane by their symbiotic bacteria (Lin et al., 2023Lin, Y.-T., Xu, T., Ip, J.C.-H., Sun, Y., Fang, L., Luan, T., Zhang, Y., Qian, P.-Y., Qiu, J.-W. (2023) Interactions among deep-sea mussels and their epibiotic and endosymbiotic chemoautotrophic bacteria: Insights from multi-omics analysis. Zoological Research 44, 106–125. https://doi.org/10.24272/j.issn.2095-8137.2022.279
), understanding the cause of the variability of La enrichment in seep mussels requires further investigation.Although the observed patterns suggest that aerobic methanotrophy-dependent mussels are typified by a distinctive accumulation of light REEs (Fig. S-3), it appears that these mussels cannot be differentiated from thiotrophy-dependent clams solely based on the extent of the La anomaly. We therefore suggest employing a (La/Nd)sn vs (Pr/Nd)sn diagram as a means of differentiation (Fig. 2; cf. Wang et al., 2020
Wang, X., Barrat, J.-A., Bayon, G., Chauvaud, L., Feng, D. (2020) Lanthanum anomalies as fingerprints of methanotrophy. Geochemical Perspectives Letters 14, 26–30. https://doi.org/10.7185/geochemlet.2019
; Barrat et al., 2022aBarrat, J.-A., Bayon, G., Carney, R.S., Chauvaud, L. (2022a) Rare earth elements as new biogeochemical proxies in deep-sea mussels. Chemical Geology 610, 121102. https://doi.org/10.1016/j.chemgeo.2022.121102
, 2023)Barrat, J.-A., Bayon, G., Lalonde, T. (2023) Calculation of cerium and lanthanum anomalies in geological and environmental samples. Chemical Geology 615, 121202. https://doi.org/10.1016/j.chemgeo.2022.121202
. In this diagram, clam soft tissues are primarily situated in the lower left corner, with (La/Nd)sn ranging from 0.93 to 2.14 and (Pr/Nd)sn ranging from 0.85 to 1.05. Similarly, clam shells occupy the same region of the diagram, with (La/Nd)sn ratios ranging from 0.99 to 1.22 and (Pr/Nd)sn ratios ranging from 0.84 to 0.91. In contrast, mussel soft tissue – (La/Nd)sn = 1.25–16.51, (Pr/Nd)sn = 0.83–2.22 – and shells – (La/Nd)sn = 3.60–8.28, (Pr/Nd)sn = 1.20–1.35 – tend toward the upper right quadrant of the diagram.Figure 2 (La/Nd)sn vs. (Pr/Nd)sn plots for (a) soft tissues and (b) shells of various macrofauna from the Haima seeps. Previous work (Bau et al., 2010
Bau, M., Balan, S., Schmidt, K., Koschinsky, A. (2010) Rare earth elements in mussel shells of the Mytilidae family as tracers for hidden and fossil high-temperature hydrothermal systems. Earth and Planetary Science Letters 299, 310–316. https://doi.org/10.1016/j.epsl.2010.09.011
; Wang et al., 2020Wang, X., Barrat, J.-A., Bayon, G., Chauvaud, L., Feng, D. (2020) Lanthanum anomalies as fingerprints of methanotrophy. Geochemical Perspectives Letters 14, 26–30. https://doi.org/10.7185/geochemlet.2019
; Barrat et al., 2022aBarrat, J.-A., Bayon, G., Carney, R.S., Chauvaud, L. (2022a) Rare earth elements as new biogeochemical proxies in deep-sea mussels. Chemical Geology 610, 121102. https://doi.org/10.1016/j.chemgeo.2022.121102
,bBarrat, J.-A., Chauvaud, L., Olivier, F., Poitevin, P., Bayon, G., Salem, D.B. (2022b) Rare earth elements and yttrium in suspension-feeding bivalves (dog cockle, Glycymeris glycymeris L.): Accumulation, vital effects and pollution. Geochimica et Cosmochimica Acta 339, 12–21. https://doi.org/10.1016/j.gca.2022.10.033
), seawater (SW; Alibo and Nozaki, 2000Alibo, D.S., Nozaki, Y. (2000) Dissolved rare earth elements in the South China Sea: geochemical characterization of the water masses. Journal of Geophysical Research 105, 28771–28783. https://doi.org/10.1029/1999JC000283
) and sediment from the Haima seeps (Wang et al., 2020Wang, X., Barrat, J.-A., Bayon, G., Chauvaud, L., Feng, D. (2020) Lanthanum anomalies as fingerprints of methanotrophy. Geochemical Perspectives Letters 14, 26–30. https://doi.org/10.7185/geochemlet.2019
) are shown for comparison.With mussels being part of the diet of the predatory gastropod Phymorhynus (Fujikura et al., 2009
Fujikura, K., Sasaki, T., Yamanaka, T., Yoshida, T. (2009) Turrids whelk, Phymorhynchus buccinoides feeds on Bathymodiolus mussels at a seep site in Sagami Bay, Japan. Plankton and Benthos Research 4, 23–30. https://doi.org/10.3800/pbr.4.23
; Sasaki et al., 2010Sasaki, T., Warén, A., Kano, Y., Okutani, T., Fujikura, K. (2010) Gastropods from recent hot vents and cold seeps: systematics, diversity and life strategies. In: Kiel, S. (Ed.) The Vent and Seep Biota. Springer, Dordrecht, 169–254. https://doi.org/10.1007/978-90-481-9572-5_7
), it is likely that the gastropod assimilates some components of the mussels, and consequently could also acquire the mussels’ La enrichment. Indeed, our findings show that the La (La/La* =1.08–3.39) and Ce anomalies (Ce/Ce* = 0.40–1.01) of P. buccinoides closely resemble those of mussels. Predation on mussels is further in accord with the (La/Nd)sn ratios from 1.02 to 3.43, and the (Pr/Nd)sn ratios from 0.88 to 1.14 found in the gills of P. buccinoides. The same argument applies to the composition of the gastropod’s visceral mass with (La/Nd)sn ratios ranging from 2.86 to 4.05 and (Pr/Nd)sn ratios ranging from 1.04 to 1.25. The shells of P. buccinoides (n = 5), with (La/Nd)sn ratios ranging from 1.04 to 1.40 and (Pr/Nd)sn ratios ranging from 0.81 to 0.89, do not exhibit significant enrichment of light REEs. Further measurements of additional predatory species with fossilisable hard parts would be needed to test whether this approach can be applied to ancient seep deposits (Kiel et al., 2016Kiel, S., Amano, K., Jenkins, R.G. (2016) Predation scar frequencies in chemosymbiotic bivalves at an Oligocene seep deposit and their potential relation to inferred sulfide tolerances. Palaeogeography, Palaeoclimatology, Palaeoecology 453, 139–145. https://doi.org/10.1016/j.palaeo.2016.04.026
).Scale worms of the genus Branchipolynoe are known to parasitise within bathymodiolin mussels (Desbruyères et al., 1985
Desbruyères, D., Gaill, F., Laubier, L., Fouquet, Y. (1985) Polychaetous annelids from hydrothermal vent ecosystems: an ecological overview. Bulletin of the Biological Society of Washington 6, 103–116.
; Becker et al., 2013Becker, E.L., Cordes, E.E., Macko, S.A., Lee, R.W., Fisher, C.R. (2013) Using stable isotope compositions of animal tissues to infer trophic interactions in Gulf of Mexico lower slope seep communities. PLoS One 8, e74459. https://doi.org/10.1371/journal.pone.0074459
), and it can therefore be anticipated that they will exhibit positive La anomalies similar to those observed in P. buccinoides. Indeed, specimens of B. pettiboneae are found to exhibit positive La anomalies (La/La* = 2.55–14.23, (La/Nd)sn = 2.18–17.42), likely due to the long term parasitic relationship between the scale worm and mussels, where the scale worm’s diet is essentially mussel based. Phymorhynchus gastropods, on the other hand, display more omnivorous feeding habits. Interestingly, the degree of the La anomaly in the scale worms is found to be even higher than that of its host mussels (Fig. 2). A potential and simple explanation is that in predator-prey relationships, metal elements become more enriched in organisms of higher trophic levels (biomagnification effect), as in the case of mercury accumulation in the food chain of aquatic organisms (Sun et al., 2020Sun, R., Yuan, J., Sonke, J.E., Zhang, Y., Zhang, T., Zheng, W., Chen, S., Meng, M., Chen, J., Liu, Y., Peng, X., Liu, C. (2020) Methylmercury produced in upper oceans accumulates in deep Mariana Trench fauna. Nature Communications 11, 3389. https://doi.org/10.1038/s41467-020-17045-3
). In summary, B. pettiboneae and some of the soft tissues of P. buccinoides (gills and visceral mass) exhibit significant enrichment of La. The corresponding La anomalies reflect the interaction between these predatory metazoan species and the aerobic methanotrophy-dependent G. haimaensis mussels within the food web structure of the Haima seep ecosystem (Fig. 3).Figure 3 Conceptual diagram of predator-prey interactions within seep ecosystems. La = lanthanum.
Finally, we put forward a Bayesian mixing model for assessing the relative contributions of various end members based on the degree of light REE enrichment, with particular focus on the La anomaly. The underlying principle of this method resembles the determination of nutritional interactions among large animals in chemosynthesis-based ecosystems using stable isotopes (Ke et al., 2022
Ke, Z., Li, R., Chen, Y., Chen, D., Chen, Z., Lian, X., Tan, Y. (2022) A preliminary study of macrofaunal communities and their carbon and nitrogen stable isotopes in the Haima cold seeps, South China Sea. Deep-Sea Research I 184, 103774. https://doi.org/10.1016/j.dsr.2022.103774
). The method is described in detail in the Supplementary Information. In short, the calculation indicates that the proportion of the La anomaly originating from the bathymodiolin mussels – and consequently the food sources – of P. buccinoides at the Haima seep site is approximately 50.4 %. This proportion is very similar to the value obtained through the estimate based on stable carbon isotopes (51.0 %; Fig. S-6). When applied to the B. pettiboneae from the Haima seep site, the proportion of La anomaly originating from the bathymodiolin mussels is approximately 87.9 %; a proportion almost identical to the value obtained with carbon isotopes (∼88.5 %; Fig. S-7). Such estimates suggest that the cycling of La and carbon between tropic levels in seep ecosystems is similar and conservative.top
Conclusions and Perspectives
Rare earth element (REE) analyses of four invertebrate taxa from the Haima seeps of the South China Sea, the thiotrophy-dependent vesicomyid clam Archivesica marissinica, the aerobic methanotrophy-dependent bathymodiolin mussel Gigantidas haimaensis, the turrid gastropod Phymorhynchus buccinoides, and the scale worm Branchipolynoe pettiboneae, revealed that REE compositions faithfully record predation on bathymodiolin mussels by the gastropod and the scale worm. By using a Bayesian mixing model, we have developed a new approach for evaluating the trophic transfer within the food chain at seeps based on REE abundances. Our results suggest that approximately 50 % of the food consumed by the gastropod is sourced from the mussels, whereas mussels could represent up to 90 % to the diet of the scale worm. The novel REE geochemistry approach presented here provides a promising tool for studying the community structure and predator-prey interactions in seep dwelling organisms.
top
Acknowledgements
Comments provided by editor Andreas Kappler and two anonymous reviewers helped to improve the manuscript. Prof. Jun Tao (Guangzhou Marine Geological Survey, China) is acknowledged for providing the image used in the graphical abstract. We thank the crew of R/V Haiyang-06 as well as the operation team of Haima ROV for their professional assistance throughout the cruises. This study was partially supported by the National Natural Science Foundation of China (Grants: 42106059 and 42225603), Shanghai Sailing Program (Grant: 21YF1416800) and Chenguang Program of Shanghai Education Development Foundation and Shanghai Municipal Education Commission (Grant: 22CGA58).
Editor: Andreas Kappler
top
References
Alibo, D.S., Nozaki, Y. (2000) Dissolved rare earth elements in the South China Sea: geochemical characterization of the water masses. Journal of Geophysical Research 105, 28771–28783. https://doi.org/10.1029/1999JC000283
 Show in context
Show in context (La/Nd)sn vs. (Pr/Nd)sn plots for (a) soft tissues and (b) shells of various macrofauna from the Haima seeps. Previous work (Bau et al., 2010; Wang et al., 2020; Barrat et al., 2022a,b), seawater (SW; Alibo and Nozaki, 2000) and sediment from the Haima seeps (Wang et al., 2020) are shown for comparison.
View in article
Barrat, J.-A., Bayon, G., Carney, R.S., Chauvaud, L. (2022a) Rare earth elements as new biogeochemical proxies in deep-sea mussels. Chemical Geology 610, 121102. https://doi.org/10.1016/j.chemgeo.2022.121102
Show in context Additionally, the La anomaly (La/La* = 1.15–2.64) observed for A. marissinica is similar to that of thiotrophy-dependent mussels from seeps on the Costa Rica margin (Barrat et al., 2022a).
View in article
The La anomalies of mussel shells (La/La* = 2.10–3.71) are similar to previous findings (La/La* = 2.50–3.92, Wang et al., 2020) – slightly higher than the La anomalies of Gigantidas shells from the Brine Pool and Bush Hill sites of the Gulf of Mexico (La/La* = 1.73–2.70), but significantly lower than the anomalies of two Bathymodiolus shells from Edison Seamount (La/La* = 7.88–11.89; Barrat et al., 2022a).
View in article
We therefore suggest employing a (La/Nd)sn vs (Pr/Nd)sn diagram as a means of differentiation (Fig. 2; cf. Wang et al., 2020; Barrat et al., 2022a, 2023).
View in article
(La/Nd)sn vs. (Pr/Nd)sn plots for (a) soft tissues and (b) shells of various macrofauna from the Haima seeps. Previous work (Bau et al., 2010; Wang et al., 2020; Barrat et al., 2022a,b), seawater (SW; Alibo and Nozaki, 2000) and sediment from the Haima seeps (Wang et al., 2020) are shown for comparison.
View in article
Barrat, J.-A., Chauvaud, L., Olivier, F., Poitevin, P., Bayon, G., Salem, D.B. (2022b) Rare earth elements and yttrium in suspension-feeding bivalves (dog cockle, Glycymeris glycymeris L.): Accumulation, vital effects and pollution. Geochimica et Cosmochimica Acta 339, 12–21. https://doi.org/10.1016/j.gca.2022.10.033
Show in context (La/Nd)sn vs. (Pr/Nd)sn plots for (a) soft tissues and (b) shells of various macrofauna from the Haima seeps. Previous work (Bau et al., 2010; Wang et al., 2020; Barrat et al., 2022a,b), seawater (SW; Alibo and Nozaki, 2000) and sediment from the Haima seeps (Wang et al., 2020) are shown for comparison.
View in article
Barrat, J.-A., Bayon, G., Lalonde, T. (2023) Calculation of cerium and lanthanum anomalies in geological and environmental samples. Chemical Geology 615, 121202. https://doi.org/10.1016/j.chemgeo.2022.121202
Show in context By determining the La and Ce anomalies (La/La* and Ce/Ce*; Eq. 1 and 2; cf. Barrat et al., 2023)
Eq. 1.
Eq. 2.
for four invertebrate species from the Haima seeps, we find that the REE patterns of the thiotrophy-dependent A. marissinica resemble the REE compositions of other seep dwelling clams (Wang et al., 2020).
View in article
We therefore suggest employing a (La/Nd)sn vs (Pr/Nd)sn diagram as a means of differentiation (Fig. 2; cf. Wang et al., 2020; Barrat et al., 2022a, 2023).
View in article
Bau, M., Balan, S., Schmidt, K., Koschinsky, A. (2010) Rare earth elements in mussel shells of the Mytilidae family as tracers for hidden and fossil high-temperature hydrothermal systems. Earth and Planetary Science Letters 299, 310–316. https://doi.org/10.1016/j.epsl.2010.09.011
Show in context (La/Nd)sn vs. (Pr/Nd)sn plots for (a) soft tissues and (b) shells of various macrofauna from the Haima seeps. Previous work (Bau et al., 2010; Wang et al., 2020; Barrat et al., 2022a,b), seawater (SW; Alibo and Nozaki, 2000) and sediment from the Haima seeps (Wang et al., 2020) are shown for comparison.
View in article
Becker, E.L., Cordes, E.E., Macko, S.A., Lee, R.W., Fisher, C.R. (2013) Using stable isotope compositions of animal tissues to infer trophic interactions in Gulf of Mexico lower slope seep communities. PLoS One 8, e74459. https://doi.org/10.1371/journal.pone.0074459
Show in context Scale worms can thrive in a wide range of habitats and frequently engage in a ‘parasitic’ relationship with mussels within seep ecosystems (Becker et al., 2013; Yao et al., 2022).
View in article
Previous stable isotope analyses conducted on scale worms from hydrocarbon seeps indicated their reliance on chemosynthesis-based biomass (Becker et al., 2013; Ke et al., 2022).
View in article
Scale worms of the genus Branchipolynoe are known to parasitise within bathymodiolin mussels (Desbruyères et al., 1985; Becker et al., 2013), and it can therefore be anticipated that they will exhibit positive La anomalies similar to those observed in P. buccinoides
View in article
Danovaro, R., Fanelli, E., Aguzzi, J., Billett, D., Carugati, L., Corinaldesi, C., Dell’Anno, A., Ggerde, K., Jamieson, A.J., Kark, S., McClain, C., Levin, L., Levin, N., Ramirez-Llodra, E., Ruhl, H., Smith, C.R., Snelgrove, P.V.R., Thomsen, L., Van Dover, C.L., Yasuhara, M. (2022) Ecological variables for developing a global deep-ocean monitoring and conservation strategy. Nature Ecology & Evolution 4, 181–192. https://doi.org/10.1038/s41559-019-1091-z
Show in context Moreover, seeps serve as nurseries for deep sea predators, which can greatly affect the diversity of animal species along continental margins (Treude et al., 2011; Danovaro et al., 2022).
View in article
Desbruyères, D., Gaill, F., Laubier, L., Fouquet, Y. (1985) Polychaetous annelids from hydrothermal vent ecosystems: an ecological overview. Bulletin of the Biological Society of Washington 6, 103–116.
Show in context Likewise, mussel fragments have been discovered among the stomach contents of the parasitic scale worm Branchipolynoe symmyitilida (Desbruyères et al., 1985).
View in article
Scale worms of the genus Branchipolynoe are known to parasitise within bathymodiolin mussels (Desbruyères et al., 1985; Becker et al., 2013), and it can therefore be anticipated that they will exhibit positive La anomalies similar to those observed in P. buccinoides
View in article
Fauchald, K., Jumars, P.A. (1979) The diet of worms: a study of polychaete feeding guilds. Oceanography and Marine Biology Annual Review 17, 193–284.
Show in context Based on their jaw structure, it has been inferred that the scale worms exhibit predatory behaviour (Fauchald and Jumars, 1979).
View in article
Fujikura, K., Sasaki, T., Yamanaka, T., Yoshida, T. (2009) Turrids whelk, Phymorhynchus buccinoides feeds on Bathymodiolus mussels at a seep site in Sagami Bay, Japan. Plankton and Benthos Research 4, 23–30. https://doi.org/10.3800/pbr.4.23
Show in context Representative examples of these higher level consumers include the turrid gastropod Phymorhynchus buccinoides and the scale worm Branchipolynoe pettiboneae at the Haima seeps of the South China Sea (Fujikura et al., 2009; Takahashi et al., 2012).
View in article
Its predation on mussels has been demonstrated by in situ behavioural observations and bait trap experiments as well as anatomic examination (Fujikura et al., 2009).
View in article
With mussels being part of the diet of the predatory gastropod Phymorhynus (Fujikura et al., 2009; Sasaki et al., 2010), it is likely that the gastropod assimilates some components of the mussels, and consequently could also acquire the mussels’ La enrichment.
View in article
Jørgensen, B.B., Boetius, A. (2007) Feast and famine — microbial life in the deep-sea bed. Nature Reviews Microbiology 5, 770–781. https://doi.org/10.1038/nrmicro1745
Show in context These ecosystems are typified by a low diversity but high abundance of the dominant species (Jørgensen and Boetius, 2007).
View in article
Ke, Z., Li, R., Chen, Y., Chen, D., Chen, Z., Lian, X., Tan, Y. (2022) A preliminary study of macrofaunal communities and their carbon and nitrogen stable isotopes in the Haima cold seeps, South China Sea. Deep-Sea Research I 184, 103774. https://doi.org/10.1016/j.dsr.2022.103774
Show in context Stable carbon and nitrogen isotope analysis conducted on P. buccinoides specimens collected at the Haima seeps indicated a dependence on local chemosynthetic carbon sources, with the gastropods taking up methane-derived carbon through predation (Ke et al., 2022).
View in article
Previous stable isotope analyses conducted on scale worms from hydrocarbon seeps indicated their reliance on chemosynthesis-based biomass (Becker et al., 2013; Ke et al., 2022).
View in article
The underlying principle of this method resembles the determination of nutritional interactions among large animals in chemosynthesis-based ecosystems using stable isotopes (Ke et al., 2022).
View in article
Kiel, S., Amano, K., Jenkins, R.G. (2016) Predation scar frequencies in chemosymbiotic bivalves at an Oligocene seep deposit and their potential relation to inferred sulfide tolerances. Palaeogeography, Palaeoclimatology, Palaeoecology 453, 139–145. https://doi.org/10.1016/j.palaeo.2016.04.026
Show in context Understanding the variables influencing the community composition of seep ecosystems, particularly the role of predation, is crucial for comprehending the succession dynamics and the evolution of chemosynthesis-based faunas in Earth history (Treude et al., 2011; Kiel et al., 2016).
View in article
Further measurements of additional predatory species with fossilisable hard parts would be needed to test whether this approach can be applied to ancient seep deposits (Kiel et al., 2016).
View in article
Levin, L.A., Baco, A.R., Bowden, D.A., Colaco, A., Cordes, E.E., Cunha, M.R., Demopoulos, A.W.J., Gobin, J., Grupe, B.M., Le, J., Metaxas, A., Netburn, A.N., Rouse, G.W., Thurber, A.R., Tunnicliffe, V., Van Dover, C.L., Vanreusel, A., Watling, L. (2016) Hydrothermal vents and methane seeps: rethinking the sphere of influence. Frontiers in Marine Science 3, 72. https://doi.org/10.3389/fmars.2016.00072
Show in context Submarine seep ecosystems are found worldwide along continental margins (Levin et al., 2016).
View in article
The impact of predation on the community structure of seep ecosystems is still not fully understood (Levin et al., 2016).
View in article
Lin, Y.-T., Xu, T., Ip, J.C.-H., Sun, Y., Fang, L., Luan, T., Zhang, Y., Qian, P.-Y., Qiu, J.-W. (2023) Interactions among deep-sea mussels and their epibiotic and endosymbiotic chemoautotrophic bacteria: Insights from multi-omics analysis. Zoological Research 44, 106–125. https://doi.org/10.24272/j.issn.2095-8137.2022.279
Show in context While the magnitude of the mussels’ La anomaly could possibly relate to the efficiency of La utilisation during aerobic oxidation of methane by their symbiotic bacteria (Lin et al., 2023), understanding the cause of the variability of La enrichment in seep mussels requires further investigation.
View in article
MacAvoy, S.E., Carney, R.S., Fisher, C.R., Macko, S.A. (2002) Use of chemosynthetic biomass by large, mobile, benthic predators in the Gulf of Mexico. Marine Ecology Progress Series 225, 65–78. https://doi.org/10.3354/meps225065
Show in context The community structure is influenced by the availability of energy sources, as well as other factors such as predation, water depth, substrate type, and ecological succession (MacAvoy et al., 2002; Morganti et al., 2022).
View in article
However, some studies have shown that predation can facilitate trophic transfer and regulate species diversity in seep ecosystems (MacAvoy et al., 2002; Olsen et al., 2014).
View in article
Morganti, T.M., Slaby, B.M., de Kluijver, A., Busch, K., Hentschel, U., Middleburg, J.J., Grotheer, H., Mollenhauer, G., Dannheim, J., Rapp, H.T., Purser, A., Boetius, A. (2022) Giant sponge grounds of Central Arctic seamounts are associated with extinct seep life. Nature Communications 13, 638. https://doi.org/10.1038/s41467-022-28129-7
Show in context The community structure is influenced by the availability of energy sources, as well as other factors such as predation, water depth, substrate type, and ecological succession (MacAvoy et al., 2002; Morganti et al., 2022).
View in article
Olsen, B.R., Troedsson, C., Hadziavdic, K., Pederson, R.B., Rapp, H.T. (2014) A molecular gut content study of Themisto abyssorum (Amphipoda) from Arctic hydrothermal vent and cold seep systems. Molecular Ecology 23, 3877–3889. https://doi.org/10.1111/mec.12511
Show in context However, some studies have shown that predation can facilitate trophic transfer and regulate species diversity in seep ecosystems (MacAvoy et al., 2002; Olsen et al., 2014).
View in article
Plum, C., Gollner, S., Martínez-Arbizu, P., Bright, M. (2015) Diversity and composition of the copepod communities associated with megafauna around a cold seep in the Gulf of Mexico with remarks on species biogeography. Marine Biodiversity 45, 419–432. https://doi.org/10.1007/s12526-014-0310-8
Show in context Predation by metazoans may also explain the differences in copepod abundance between mussel beds and tubeworm colonies at seeps in the Gulf of Mexico (Plum et al., 2015).
View in article
Portail, M., Olu, K., Dubois, S.F., Escobar-Briones, E., Gelinas, Y., Menot, L., Sarrazin, J. (2016) Food-web complexity in Guaymas Basin hydrothermal vents and cold seeps. PLoS One 11, e0162263. https://doi.org/10.1371/journal.pone.0162263
Show in context Previous studies have suggested that predators in these ecosystems do not play a dominate role and their influence is typically considered modest (Portail et al., 2016).
View in article
Regarding their trophic level, mussels can be classified as primary consumers (Portail et al., 2016).
View in article
Pourmand, A., Dauphas, N., Ireland, T.J. (2012) A novel extraction chromatography and MC-ICP-MS technique for rapid analysis of REE, Sc and Y: Revising CI-chondrite and Post-Archean Australian Shale (PAAS) abundances. Chemical Geology 291, 38–54. https://doi.org/10.1016/j.chemgeo.2011.08.011
Sasaki, T., Warén, A., Kano, Y., Okutani, T., Fujikura, K. (2010) Gastropods from recent hot vents and cold seeps: systematics, diversity and life strategies. In: Kiel, S. (Ed.) The Vent and Seep Biota. Springer, Dordrecht, 169–254. https://doi.org/10.1007/978-90-481-9572-5_7
Show in context Turrid gastropods of the genus Phymorhynchus are common in various deep sea habitats such as hydrothermal vents, hydrocarbon seeps, and sunken wood, where they are unspecialised predators that benefit from the high mass of available food in these ecosystems (Sasaki et al., 2010).
View in article
Phymorhynchus has a well developed, funnel-shaped rhynchostome that facilitates its predatory behaviour (Warén and Bouchet, 2001), and fragments of various organisms in the gut of Phymorhynchus have been documented (Sasaki et al., 2010).
View in article
With mussels being part of the diet of the predatory gastropod Phymorhynus (Fujikura et al., 2009; Sasaki et al., 2010), it is likely that the gastropod assimilates some components of the mussels, and consequently could also acquire the mussels’ La enrichment.
View in article
Semrau, J.D., Dispirito, A.A., Gu, W., Yoon, S. (2018) Metals and methanotrophy. Applied and Environmental Microbiology 84, e02289–17. https://doi.org/10.1128/AEM.02289-17
Show in context This approach is based on the facts that (1) the occurrence of La anomalies in seep dwelling metazoans is a robust fingerprint of methanotrophy since the second step in the aerobic oxidation of methane (CH3OH → HCHO) can be catalysed by La (Semrau et al., 2018; Wang et al., 2020), and (2) aerobic methanotrophy-dependent mussels are ubiquitous in seep ecosystems (Wang et al., 2022).
View in article
Sun, R., Yuan, J., Sonke, J.E., Zhang, Y., Zhang, T., Zheng, W., Chen, S., Meng, M., Chen, J., Liu, Y., Peng, X., Liu, C. (2020) Methylmercury produced in upper oceans accumulates in deep Mariana Trench fauna. Nature Communications 11, 3389. https://doi.org/10.1038/s41467-020-17045-3
Show in context A potential and simple explanation is that in predator-prey relationships, metal elements become more enriched in organisms of higher trophic levels (biomagnification effect), as in the case of mercury accumulation in the food chain of aquatic organisms (Sun et al., 2020).
View in article
Takahashi, Y., Sasaki, Y., Chikaraishi, Y., Tsuchiya, M., Watanabe, H., Asahida, T., Maruyama, T., Fujikura, K. (2012) Does the symbiotic scale-worm feed on the host mussel in deep-sea vent fields? Researches in Organic Geochemistry 28, 23–26.
Show in context Representative examples of these higher level consumers include the turrid gastropod Phymorhynchus buccinoides and the scale worm Branchipolynoe pettiboneae at the Haima seeps of the South China Sea (Fujikura et al., 2009; Takahashi et al., 2012).
View in article
Nitrogen stable isotope compositions of amino acids of the scale worm B. pettiboneae and its host mussel indicate that the mussel serves as the primary source of amino acids for the adult parasitic worm (Takahashi et al., 2012).
View in article
Treude, T., Kiel, S., Linke, P., Peckmann, J., Goedert, J.L. (2011) Elasmobranch egg capsules associated with modern and ancient cold seeps: a nursery for marine deep-water predators. Marine Ecology Progress Series 437, 175–181. https://doi.org/10.3354/meps09305
Show in context Moreover, seeps serve as nurseries for deep sea predators, which can greatly affect the diversity of animal species along continental margins (Treude et al., 2011; Danovaro et al., 2022).
View in article
Understanding the variables influencing the community composition of seep ecosystems, particularly the role of predation, is crucial for comprehending the succession dynamics and the evolution of chemosynthesis-based faunas in Earth history (Treude et al., 2011; Kiel et al., 2016).
View in article
van Denderen, P.D., Lindegren, M., MacKenzie, B.R., Watson, R.A., Andersen, K.H. (2018) Global patterns in marine predatory fish. Nature Ecology & Evolution 2, 65–70. https://doi.org/10.1038/s41559-017-0388-z
Show in context Overall, predator-prey relationships are recognised as important mechanisms in structuring ecosystems (van Denderen et al., 2018).
View in article
Van Gaever, S., Galeron, J., Sibuet, M., Vanreusel, A. (2009) Deep-sea habitat heterogeneity influence on meiofaunal communities in the Gulf of Guinea. Deep-Sea Research II 56, 2259–2269. https://doi.org/10.1016/j.dsr2.2009.04.008
Show in context For example, predation pressure has been found to be the main factor behind the inverse correlation between macrofauna and meiofauna/nematode densities among the seep benthos (Van Gaever et al., 2009).
View in article
Wang, X., Barrat, J.-A., Bayon, G., Chauvaud, L., Feng, D. (2020) Lanthanum anomalies as fingerprints of methanotrophy. Geochemical Perspectives Letters 14, 26–30. https://doi.org/10.7185/geochemlet.2019
Show in context This approach is based on the facts that (1) the occurrence of La anomalies in seep dwelling metazoans is a robust fingerprint of methanotrophy since the second step in the aerobic oxidation of methane (CH3OH → HCHO) can be catalysed by La (Semrau et al., 2018; Wang et al., 2020), and (2) aerobic methanotrophy-dependent mussels are ubiquitous in seep ecosystems (Wang et al., 2022).
View in article
By determining the La and Ce anomalies (La/La* and Ce/Ce*; Eq. 1 and 2; cf. Barrat et al., 2023)
Eq. 1.
Eq. 2.
for four invertebrate species from the Haima seeps, we find that the REE patterns of the thiotrophy-dependent A. marissinica resemble the REE compositions of other seep dwelling clams (Wang et al., 2020).
View in article
The La anomalies of mussel shells (La/La* = 2.10–3.71) are similar to previous findings (La/La* = 2.50–3.92, Wang et al., 2020) – slightly higher than the La anomalies of Gigantidas shells from the Brine Pool and Bush Hill sites of the Gulf of Mexico (La/La* = 1.73–2.70), but significantly lower than the anomalies of two Bathymodiolus shells from Edison Seamount (La/La* = 7.88–11.89; Barrat et al., 2022a).
View in article
We therefore suggest employing a (La/Nd)sn vs (Pr/Nd)sn diagram as a means of differentiation (Fig. 2; cf. Wang et al., 2020; Barrat et al., 2022a, 2023).
View in article
(La/Nd)sn vs. (Pr/Nd)sn plots for (a) soft tissues and (b) shells of various macrofauna from the Haima seeps. Previous work (Bau et al., 2010; Wang et al., 2020; Barrat et al., 2022a,b), seawater (SW; Alibo and Nozaki, 2000) and sediment from the Haima seeps (Wang et al., 2020) are shown for comparison.
View in article
(La/Nd)sn vs. (Pr/Nd)sn plots for (a) soft tissues and (b) shells of various macrofauna from the Haima seeps. Previous work (Bau et al., 2010; Wang et al., 2020; Barrat et al., 2022a,b), seawater (SW; Alibo and Nozaki, 2000) and sediment from the Haima seeps (Wang et al., 2020) are shown for comparison.
View in article
Wang, X., Guan, H., Qiu, J.-W., Xu, T., Peckmann, J., Chen, D., Feng, D. (2022) Macro-ecology of cold seeps in the South China Sea. Geosystems and Geoenvironment 1, 100081. https://doi.org/10.1016/j.geogeo.2022.100081
Show in context This approach is based on the facts that (1) the occurrence of La anomalies in seep dwelling metazoans is a robust fingerprint of methanotrophy since the second step in the aerobic oxidation of methane (CH3OH → HCHO) can be catalysed by La (Semrau et al., 2018; Wang et al., 2020), and (2) aerobic methanotrophy-dependent mussels are ubiquitous in seep ecosystems (Wang et al., 2022).
View in article
Warén, A., Bouchet, P. (2001) Gastropoda and Monoplacophora from hydrothermal vents and seeps; new taxa and records. The Veliger 44, 116–231.
Show in context Phymorhynchus has a well developed, funnel-shaped rhynchostome that facilitates its predatory behaviour (Warén and Bouchet, 2001), and fragments of various organisms in the gut of Phymorhynchus have been documented (Sasaki et al., 2010).
View in article
Yao, G., Zhang, H., Xiong, P., Jia, H., He, M. (2022) Effects of scale worm parasitism on interactions between the symbiotic gill microbiome and gene regulation in deep sea mussel hosts. Frontiers in Microbiology 13, 940766. https://doi.org/10.3389/fmicb.2022.940766
Show in context Scale worms can thrive in a wide range of habitats and frequently engage in a ‘parasitic’ relationship with mussels within seep ecosystems (Becker et al., 2013; Yao et al., 2022).
View in article
Parasitism of B. pettiboneae within G. haimaensis was confirmed and described in detail by Yao et al. (2022).
View in article
top
Supplementary Information
The Supplementary Information includes:
- Materials and Methods
- Tables S-1 to S-7
- Figures S-1 to S-7
- Supplementary Information References
Download the Supplementary Information (PDF)