



Molecular biosignatures in planetary analogue salts: implications for transport of organics in sulfate-rich brines beyond Earth
Affiliations | Corresponding Author | Cite as | Funding information
Published by the European Association of Geochemistry
under Creative Commons License CC BY 4.0
Keywords: Astrobiology, organics, evolved gas analysis, salts, sulfate-rich brines, transport

- Share this article




-
Article views:476Cumulative count of HTML views and PDF downloads.
- Download Citation
- Rights & Permissions

top
Abstract
Figures and Tables
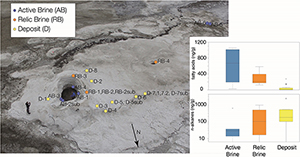
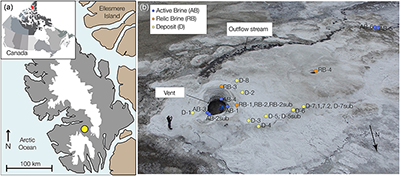 Figure 1 (a) Lost Hammer salt deposit location (yellow dot), Axel Heiberg Island. (b) Lost Hammer overview with geologist for scale. Sampling points are marked by coloured dots indicating the 3 different areas into which the Lost Hammer locality was divided: Active Brine, Relic Brine, and Deposit. Sample names ending in –sub indicate locations where a subsurface sample was paired with a corresponding surface salt. | 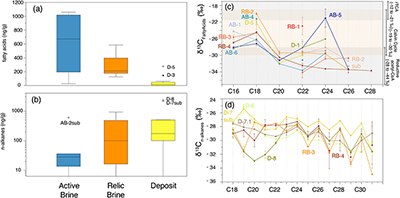 Figure 2 Whisker plot for (a) total FA and (b) total n-alkanes, concentrations in ng/g. The dots are outlier samples. Individual distributions can be found in Figures S-1, S-2 and S-3. Carbon isotopes from n-fatty acid compounds (c) and n-alkanes (d). The δ13C values for FA range between –21 ± 2.17 ‰ to –34 ± 0.01 ‰. The δ13C n-alkanes range between –25 ± 0.4 ‰ and –34 ± 0.01 ‰; complete data set in Table S-3. | 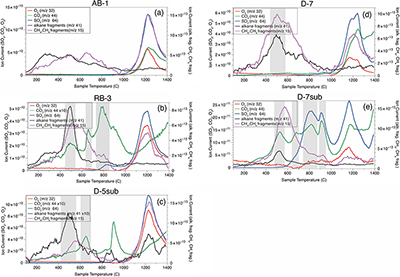 Figure 3 Subset representing measured samples for EGA-MS, complete sample set in Figure S-4. (a) Active Brine sample AB-1, (b) Relic Brine sample RB-3, (c) Deposit sample D-5sub, (d) D-7.1 and (e) D-7sub. All samples evolve SO2 (m/z 64), O2 (m/z 32), and CO2 (m/z 44) during heating at temperatures up to 1400 °C. All samples apart from RB-3 show the evolution of likely alkane fragments (e.g., signal at m/z 41) and possible methane (either discrete methane or CH3 fragments of a larger organic molecule; signal at m/z 15; see SI). Grey area indicates peaks of interest discussed in text. | 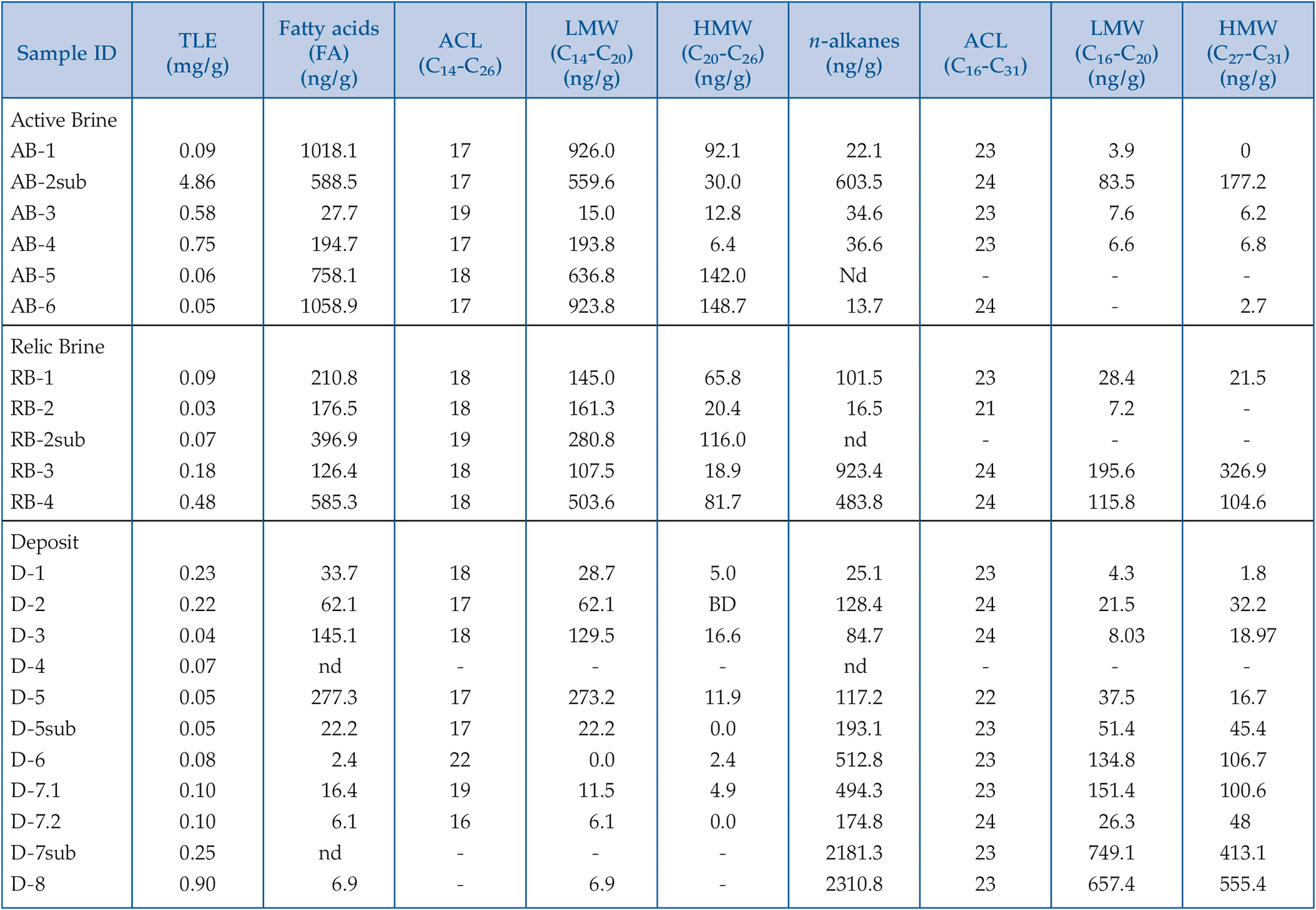 Table 1 Organic biomarker results for FA and n-alkanes. Total Lipid Extract (TLE). Average Chain Length (ACL) for n-fatty acids and n-alkanes calculated following equations from Carrizo et al. (2019). FA (branched and non-branched) and n-alkane concentrations are split into concentrations of Low Molecular Weight (LMW) and High Molecular Weight (HMW) compounds. Non-detected as ‘nd’, equations with non-detected carbon chains as ‘-’, below detection as “BD”.Complete data set in Table S-1 and S-2 for FA and for n-alkanes. |
| Figure 1 | Figure 2 | Figure 3 | Table 1 |
top
Introduction
Salt minerals precipitated during evaporation or freezing of brines can capture organic and geochemical biosignatures, preserving crucial information about the aqueous environment at the time of their formation (Schopf et al., 2012
Schopf, J.W., Farmer, J.D., Foster, I.S., Kudryavtsev, A.B., Gallardo, V.A., Espinoza, C. (2012) Gypsum-permineralized microfossils and their relevance to the search for life on Mars. Astrobiology 12, 619–633. https://doi.org/10.1089/ast.2012.0827
). Such salts are prevalent throughout the Solar System, including Mars, icy moons, and asteroids. Their association with liquid water environments make salts high priority astrobiology targets (Phillips et al., 2023Phillips, M.S., McInenly, M., Hofmann, M.H., Hinman, N.W., Warren-Rhodes, K., et al. (2023) Salt Constructs in Paleo-Lake Basins as High-Priority Astrobiology Targets. Remote Sensing 15, 314. https://doi.org/10.3390/rs15020314
). On Mars, sulfate salts are abundant globally and record major shifts in aqueous environment chemistry (Bibring et al., 2006Bibring, J.P., Langevin, Y., Mustard, J.F., Poulet, F., Arvidson, R., et al. (2006) Global mineralogical and aqueous Mars history derived from OMEGA/Mars Express data. Science 312, 400–404. https://doi.org/10.1126/science.1122659
). Hydrated chloride and sulfate salts are suggested to be present on the surface of Europa (Brown and Hand, 2013Brown, M.E., Hand, K.P. (2013) Salts and radiation products on the surface of Europa. The Astronomical Journal 145, 110. https://doi.org/10.1088/0004-6256/145/4/110
), originating from a range of possible endogenous and/or exogenous processes, including emplacement from subsurface liquid reservoirs (King et al., 2022King, O., Fletcher, L.N., Ligier, N. (2022) Compositional mapping of Europa using MCMC modeling of near-IR VLT/SPHERE and Galileo/NIMS observations. The Planetary Science Journal 3, 72. https://doi.org/10.3847/PSJ/ac596d
). Chloride and carbonate salts sourced from subsurface water-rock reactions are components of ice grains in Enceladus plumes (Postberg et al., 2009Postberg, F., Kempf, S., Schmidt, J., Brilliantov, N., Beinsen, A., et al. (2009) Sodium salts in E-ring ice grains from an ocean below the surface of Enceladus. Nature 459, 1098–1101. https://doi.org/10.1038/nature08046
), and carbonate and chloride salts are present on dwarf planet Ceres, likely sourced from a liquid layer beneath the crust (De Sanctis et al., 2020De Sanctis, M.C., Ammannito, E., Raponi, A., Frigeri, A., Ferrari, M., et al. (2020) Fresh emplacement of hydrated sodium chloride on Ceres from ascending salty fluids. Nature Astronomy 4, 786–793. https://doi.org/10.1038/s41550-020-1138-8
). Across all these targets, salt minerals have the potential to capture evidence of putative past or present microbial activity, in addition to abiogenic organic matter (Chan et al., 2018Chan, Q.H., Zolensky, M.E., Kebukawa, Y., Fries, M., Ito, M., et al. (2018) Organic matter in extraterrestrial water-bearing salt crystals. Science Advances 4, eaao3521. https://doi.org/10.1126/sciadv.aao3521
).Terrestrial environments containing hydrated and polyhydrated salts, similar to those found in extraterrestrial deposits can provide a framework for understanding the capture and detection of microbial organic biosignatures. Organic biosignatures, such as lipids, are fundamental components of cell structures and are utilised as molecular markers to identify microbial activity, environmental processes, and their stable carbon isotopic ratios can reveal autotrophic carbon fixation pathways (Jahnke et al., 2019
Jahnke, L.L., Des Marais, D.J. (2019) Carbon isotopic composition of lipid biomarkers from an endoevaporitic gypsum crust microbial mat reveals cycling of mineralized organic carbon. Geobiology 17, 643–659. https://doi.org/10.1111/gbi.12355
).The analysis of materials from relevant planetary analogue environments with flight-like techniques can further support the interpretation of data from flight analyses of planetary materials. Evolved gas analysis mass spectrometry (EGA-MS) studies of salt-bearing planetary materials are currently being carried out by the Sample Analysis at Mars (SAM) instrument suite on the Mars Science Laboratory Curiosity rover (e.g., Eigenbrode et al., 2018
Eigenbrode, J.L., Summons, R.E., Steele, A., Freissinet, C., Millan, M., et al. (2018) Organic matter preserved in 3-billion-year-old mudstones at Gale crater, Mars. Science 360, 1096–1101. https://doi.org/10.1126/science.aas9185
). Similar thermal analyses are planned for future in situ planetary missions (Reinhardt et al., 2020Reinhardt, M., Goetz, W., Thiel, V. (2020) Testing flight-like pyrolysis gas chromatography–mass spectrometry as performed by the Mars Organic Molecule Analyzer onboard the ExoMars 2020 rover on Oxia Planum analog samples. Astrobiology 20, 415–428. https://doi.org/10.1089/ast.2019.2143
). The significance of this research lines in the combination of EGA-MS and molecular results, which have implications for past, current, and future in situ analyses on sulfate salts found throughout the Solar System.top
Study Area
Axel Heiberg Island (Canadian High Arctic) hosts perennially cold hypersaline springs linked to sub-permafrost evaporite diapirs (Pollard et al., 1999
Pollard, W., Omelon, C., Andersen, D., McKay, C. (1999) Perennial spring occurrence in the Expedition Fiord area of western Axel Heiberg Island, Canadian high Arctic. Canadian Journal of Earth Sciences 36, 105–120. https://doi.org/10.1139/e98-097
). The Lost Hammer (LH) spring (Fig. 1a) possesses the lowest temperature (−5 °C) and highest concentration of salt (24 %) of these springs, and releases gases dominated by methane (50 %) (Pollard et al., 1999Pollard, W., Omelon, C., Andersen, D., McKay, C. (1999) Perennial spring occurrence in the Expedition Fiord area of western Axel Heiberg Island, Canadian high Arctic. Canadian Journal of Earth Sciences 36, 105–120. https://doi.org/10.1139/e98-097
). During sampling of the LH spring in July 2017, the LH outflow brine originated from a single observable source comprising a salt dome with a ∼2 m diameter central outlet (hereafter termed ‘vent’; Fig. 1b). Adjacent to the vent dome, a ∼1 m high salt apron extends ∼100 m downslope, with a maximum width of ∼50 m (hereafter termed ‘apron deposit’; Fig. 1b). A dry cavity (∼1 m diameter) in the apron deposit and associated dry channel indicated the likely location of past brine flow activity (hereafter referred to as ‘relic brine’; Fig. 1b).Figure 1 (a) Lost Hammer salt deposit location (yellow dot), Axel Heiberg Island. (b) Lost Hammer overview with geologist for scale. Sampling points are marked by coloured dots indicating the 3 different areas into which the Lost Hammer locality was divided: Active Brine, Relic Brine, and Deposit. Sample names ending in –sub indicate locations where a subsurface sample was paired with a corresponding surface salt.
At time of sampling, the vent dome and apron deposit comprised sodium sulfates and chlorides of varying hydration states, with the highest abundances of sulfates closest to the vent (Fox-Powell et al., 2019
Fox‐Powell, M.G., Osinski, G.R., Applin, D., Stromberg, J.M., Gázquez, F., et al. (2019) Natural analogue constraints on Europa’s non‐ice surface material. Geophysical Research Letters 46, 5759–5767. https://doi.org/10.1029/2018GL081339
). Active microbial communities in LH vent and outflow stream are characterised by low concentrations of biomass in the sediments (around 105 cells/g) and lithotrophic microorganisms adapted to hypersaline and cold conditions: anaerobic methane oxidising archaea and sulfate reducing/sulfide oxidising bacteria are prevalent (Lay et al., 2013Lay, C.Y., Mykytczuk, N.C., Yergeau, E., Lamarche-Gagnon, G., Greer, C.W., Whyte, L.G. (2013) Defining the functional potential and active community members of a sediment microbial community in a high-arctic hypersaline subzero spring. Applied and Environmental Microbiology 79, 3637–3648. https://doi.org/10.1128/AEM.00153-13
; Lamarche-Gagnon et al., 2015Lamarche-Gagnon, G., Comery, R., Greer, C.W., Whyte, L.G. (2015) Evidence of in situ microbial activity and sulphidogenesis in perennially sub-0°C and hypersaline sediments of a high Arctic permafrost spring. Extremophiles 19, 1–15. https://doi.org/10.1007/s00792-014-0703-4
; Sapers et al., 2017Sapers, H.M., Ronholm, J., Raymond-Bouchard, I., Comrey, R., Osinski, G.R., Whyte, L.G. (2017) Biological characterization of microenvironments in a hypersaline cold spring Mars analog. Frontiers in Microbiology 8, 2527.https://doi.org/10.3389/fmicb.2017.02527
; Magnuson et al., 2022Magnuson, E., Altshuler, I., Fernández-Martínez, M.Á., Chen, Y.J., Maggiori, C., et al. (2022) Active lithoautotrophic and methane-oxidizing microbial community in an anoxic, sub-zero, and hypersaline High Arctic spring. The ISME Journal 16, 1798–1808. https://doi.org/10.1038/s41396-022-01233-8
). Quadruple Sulfur Isotope (δ34S, Δ33S and Δ36S) analysis of iron sulfide from the outflow brine support these observations with fractionations typical of microbial sulfate reduction and sulfur disproportionation (Moreras-Marti et al., 2021Moreras-Marti, A., Fox-Powell, M., Stueeken, E., Di Rocco, T., Galloway, T., et al. (2021) Quadruple sulfur isotope biosignatures from terrestrial Mars analogue systems. Geochimica et Cosmochimica Acta 308, 157–172. https://doi.org/10.1016/j.gca.2021.06.007
).top
Methods
Between 15 to 40 g of salt were collected for each sample in polyethylene bags from the LH deposit and maintained at 4 °C as described in Fox-Powell et al. (2019)
Fox‐Powell, M.G., Osinski, G.R., Applin, D., Stromberg, J.M., Gázquez, F., et al. (2019) Natural analogue constraints on Europa’s non‐ice surface material. Geophysical Research Letters 46, 5759–5767. https://doi.org/10.1029/2018GL081339
. Samples were divided based on criteria of area: (1) Active Brine (AB) - within the active brine flow or source vent, (2) Relic Brine (RB) - samples on the outer edge of the vent and from the secondary dry vent and channel structure desribed above, but with no contact with actively liquid brine flow, and (3) Deposit (D) - older salt apron, with no contact with liquid brine channel. Samples D-7.1 and D-7.2 are two replicates from the same Deposit parent sample. Samples at depth (‘sub’) were collected at ∼4 cm from the surface. Samples were freeze dried for organic biomarker extraction and analyses and EGA-MS; details are provided in Supplementary Information.top
Results and Discussion
Sources of fatty acids and n-alkanes. Fatty acids (FA) exhibited a biological signature throughout the LH salt deposit (Fig. 2a,c). This was evidenced by the presence of short-to-mid chain FA (C12-C24) of even-over-odd preference, and Cmax at C16 and C18, the dominant FA lengths for bacteria and eukaryotic cells (López-Lara and Geiger, 2010
López-Lara, I.M., Geiger, O. (2010) Formation of fatty acids. In: Timmis, K.N. (Ed.) Handbook of Hydrocarbon and Lipid Microbiology. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-540-77587-4_26
). The greatest range in FA concentrations were observed in Active Brine salts (27.7 to 1072.5 ng/g; Fig. 2a), followed by Relic Brine salts (126.4 to 483.7 ng/g). The lowest concentration and range were observed in Deposit salts (6.9 to 277.3 ng/g). Several samples across the Active Brine and Relic Brine salts yielded branched FA (i- for isomer, a for anti-isomer). The i-C15, a-C15, i-C17 and a-C17 are interpreted to be associated with microbial sulfate reducers (MSR), while C18:1 records the presence of cyanobacteria or other types of bacteria and algae (Perry et al., 1979Perry, G.J., Volkman, J.K., Johns, R.B. (1979) Fatty Acids of Bacterial Origin in Contemporary Marine Sediments. Geochimica et Cosmochimica Acta 43, 1715–1725. https://doi.org/10.1016/0016-7037(79)90020-6
). Such branched FA have been identified in hypersaline lakes (Nichols et al., 2023Nichols, F., Pontefract, A., Dion‐Kirschner, H., Masterson, A.L., Osburn, M.R. (2023) Lipid Biosignatures From SO4‐Rich Hypersaline Lakes of the Cariboo Plateau. Journal of Geophysical Research: Biogeosciences 128, e2023JG007480 https://doi.org/10.1029/2023JG007480
). Unsaturated i-C15 and a-C15 could not be identified as compounds under C16 had become degraded before measuring for branched and unsaturated FA. The presence of i-C17, a-C17 and C18:1 at the LH Deposit corroborates past studies of sulfur isotopes and microbial analyses which identified microbial sulfate reduction and sulfur oxidation occurring in the streams and vent sediments (Moreras-Marti et al, 2021Moreras-Marti, A., Fox-Powell, M., Stueeken, E., Di Rocco, T., Galloway, T., et al. (2021) Quadruple sulfur isotope biosignatures from terrestrial Mars analogue systems. Geochimica et Cosmochimica Acta 308, 157–172. https://doi.org/10.1016/j.gca.2021.06.007
; Magnuson et al., 2022Magnuson, E., Altshuler, I., Fernández-Martínez, M.Á., Chen, Y.J., Maggiori, C., et al. (2022) Active lithoautotrophic and methane-oxidizing microbial community in an anoxic, sub-zero, and hypersaline High Arctic spring. The ISME Journal 16, 1798–1808. https://doi.org/10.1038/s41396-022-01233-8
). Furthermore, the C isotope results from LH FA (C16 and C18 from −15 to −28 ‰ and >C20 from –21 to −34 ‰) (Fig. 2 c,d), fall in the ranges for autotrophic pathways with a mixed use of three assimilation pathways of carbon including: Reverse tricarboxylic acid (rTCA) cycle for autotrophic bacteria (from −12 to −21 ‰), Calvin cycle for cyanobacteria and algae (from –19 to −30 ‰) and reductive acetyl-CoA pathway for anaerobic bacteria and archaea (−28 to −44 ‰) (Jahnke et al., 2019Jahnke, L.L., Des Marais, D.J. (2019) Carbon isotopic composition of lipid biomarkers from an endoevaporitic gypsum crust microbial mat reveals cycling of mineralized organic carbon. Geobiology 17, 643–659. https://doi.org/10.1111/gbi.12355
). The rapid degradation of FA through biological processes, occurring as quickly as a few weeks, underscores their presence in terrestrial environments as potential indicators of recent microbial activity (Perry et al., 1979Perry, G.J., Volkman, J.K., Johns, R.B. (1979) Fatty Acids of Bacterial Origin in Contemporary Marine Sediments. Geochimica et Cosmochimica Acta 43, 1715–1725. https://doi.org/10.1016/0016-7037(79)90020-6
), such as observed in Active Brine samples, and to a lesser extent in Relic Brine samples. The limited occurrence of FA in Deposit samples is consistent with the absence of recent microbial activity and suggests rapid degradation of deposited compounds.Figure 2 Whisker plot for (a) total FA and (b) total n-alkanes, concentrations in ng/g. The dots are outlier samples. Individual distributions can be found in Figures S-1, S-2 and S-3. Carbon isotopes from n-fatty acid compounds (c) and n-alkanes (d). The δ13C values for FA range between –21 ± 2.17 ‰ to –34 ± 0.01 ‰. The δ13C n-alkanes range between –25 ± 0.4 ‰ and –34 ± 0.01 ‰; complete data set in Table S-3.
The n-alkanes and carbon isotope values indicate various biological sources in LH. The general prevalence of Medium Molecular Weight (MMW; C21 to C26) over High Molecular Weight (HMW; >C27) and Low Molecular Weight (LMW; <C20) n-alkanes (Table 1) is indicative of algal and/or cyanobacteria sources (Castañeda and Shouten (2011)
Castañeda, I.S., Schouten, S. (2011) A review of molecular organic proxies for examining modern and ancient lacustrine environments. Quaternary Science Reviews 30, 2851–2891. https://doi.org/10.1016/j.quascirev.2011.07.009.
, and references therein). The highest concentrations of n-alkanes (C16 to C32) were observed in Deposit samples (13.7 to 2310.8 ng/g), followed by Relic Brine (16.48 to 923.4 ng/g) and lowest in Active Brine (34.6 to 603.5 ng/g) (Fig. 2b,c, Table 1). General odd/even dominance of HMW n-alkanes pointed to the contribution of higher plants and/or lichens which predominantely produce HMW n-alkanes (Castañeda and Shouten, 2011Castañeda, I.S., Schouten, S. (2011) A review of molecular organic proxies for examining modern and ancient lacustrine environments. Quaternary Science Reviews 30, 2851–2891. https://doi.org/10.1016/j.quascirev.2011.07.009.
). Likewise samples RB-3, RB-4 and D-8 present n-alkane Cmax peaks typical of higher plants as HMW n-alkanes are known to be major components of waxes in vascular land plants (Eglinton and Hamilton, 1967Eglinton, G., Hamilton, R.J. (1967) Leaf Epicuticular Waxes: The waxy outer surfaces of most plants display a wide diversity of fine structure and chemical constituents. Science 156, 1322–1335. https://doi.org/10.wa1126/science.156.3780.1322
). No visible plant growth was observed within the outflow stream or LH salts themselves, so these are likely sourced from local tundra vegetation nearby LH or transported by aeolian processes. The LMW n-alkane distribution for all samples had no odd/even carbon number preference, suggesting derivation from microbial lipids, and/or microbial re-working of plant n-alkanes through early diagenesis (Castañeda and Shouten, 2011Castañeda, I.S., Schouten, S. (2011) A review of molecular organic proxies for examining modern and ancient lacustrine environments. Quaternary Science Reviews 30, 2851–2891. https://doi.org/10.1016/j.quascirev.2011.07.009.
). The δ13C of LMW n-alkanes varied between −27 and −32 ‰, the MMW −27 and −32 ‰ and HMW −28 and −34 ‰, suggesting an origin from bacteria, algae and or phytoplankton. The HMW n-alkanes δ13C also suggested an input from C3 plants which usually exhibit a δ13C of −34 to −35 ‰, (Castañeda and Shouten, 2011Castañeda, I.S., Schouten, S. (2011) A review of molecular organic proxies for examining modern and ancient lacustrine environments. Quaternary Science Reviews 30, 2851–2891. https://doi.org/10.1016/j.quascirev.2011.07.009.
).Table 1 Organic biomarker results for FA and n-alkanes. Total Lipid Extract (TLE). Average Chain Length (ACL) for n-fatty acids and n-alkanes calculated following equations from Carrizo et al. (2019)
Carrizo, D., Sánchez-García, L., Rodriguez, N., Gómez, F. (2019) Lipid biomarker and carbon stable isotope survey on the Dallol hydrothermal system in Ethiopia. Astrobiology 19, 1474–1489. https://doi.org/10.1089/ast.2018.1963
. FA (branched and non-branched) and n-alkane concentrations are split into concentrations of Low Molecular Weight (LMW) and High Molecular Weight (HMW) compounds. Non-detected as ‘nd’, equations with non-detected carbon chains as ‘-’, below detection as “BD”.Complete data set in Table S-1 and S-2 for FA and for n-alkanes.| Sample ID | TLE (mg/g) | Fatty acids(FA) (ng/g) | ACL (C14-C26) | LMW (C14-C20) (ng/g) | HMW (C20-C26) (ng/g) | n-alkanes (ng/g) | ACL (C16-C31) | LMW (C16-C20) (ng/g) | HMW (C27-C31) (ng/g) |
| Active Brine | |||||||||
| AB-1 | 0.09 | 1018.1 | 17 | 926.0 | 92.1 | 22.1 | 23 | 3.9 | 0 |
| AB-2sub | 4.86 | 588.5 | 17 | 559.6 | 30.0 | 603.5 | 24 | 83.5 | 177.2 |
| AB-3 | 0.58 | 27.7 | 19 | 15.0 | 12.8 | 34.6 | 23 | 7.6 | 6.2 |
| AB-4 | 0.75 | 194.7 | 17 | 193.8 | 6.4 | 36.6 | 23 | 6.6 | 6.8 |
| AB-5 | 0.06 | 758.1 | 18 | 636.8 | 142.0 | Nd | - | - | - |
| AB-6 | 0.05 | 1058.9 | 17 | 923.8 | 148.7 | 13.7 | 24 | - | 2.7 |
| Relic Brine | |||||||||
| RB-1 | 0.09 | 210.8 | 18 | 145.0 | 65.8 | 101.5 | 23 | 28.4 | 21.5 |
| RB-2 | 0.03 | 176.5 | 18 | 161.3 | 20.4 | 16.5 | 21 | 7.2 | - |
| RB-2sub | 0.07 | 396.9 | 19 | 280.8 | 116.0 | nd | - | - | - |
| RB-3 | 0.18 | 126.4 | 18 | 107.5 | 18.9 | 923.4 | 24 | 195.6 | 326.9 |
| RB-4 | 0.48 | 585.3 | 18 | 503.6 | 81.7 | 483.8 | 24 | 115.8 | 104.6 |
| Deposit | |||||||||
| D-1 | 0.23 | 33.7 | 18 | 28.7 | 5.0 | 25.1 | 23 | 4.3 | 1.8 |
| D-2 | 0.22 | 62.1 | 17 | 62.1 | BD | 128.4 | 24 | 21.5 | 32.2 |
| D-3 | 0.04 | 145.1 | 18 | 129.5 | 16.6 | 84.7 | 24 | 8.03 | 18.97 |
| D-4 | 0.07 | nd | - | - | - | nd | - | - | - |
| D-5 | 0.05 | 277.3 | 17 | 273.2 | 11.9 | 117.2 | 22 | 37.5 | 16.7 |
| D-5sub | 0.05 | 22.2 | 17 | 22.2 | 0.0 | 193.1 | 23 | 51.4 | 45.4 |
| D-6 | 0.08 | 2.4 | 22 | 0.0 | 2.4 | 512.8 | 23 | 134.8 | 106.7 |
| D-7.1 | 0.10 | 16.4 | 19 | 11.5 | 4.9 | 494.3 | 23 | 151.4 | 100.6 |
| D-7.2 | 0.10 | 6.1 | 16 | 6.1 | 0.0 | 174.8 | 24 | 26.3 | 48 |
| D-7sub | 0.25 | nd | - | - | - | 2181.3 | 23 | 749.1 | 413.1 |
| D-8 | 0.90 | 6.9 | - | 6.9 | - | 2310.8 | 23 | 657.4 | 555.4 |
Association of molecular biosignatures with sulfate salts. A subset of seven representative samples capturing the full spatial distribution of the LH salt deposit were selected based on available remaining sample material for EGA-MS (Figs. 3, S-3). The EGA-MS results from Active Brine samples indicate CO2, CH4 or CH3- fragments of molecules evolving coincident with Na sulfate thermal decomposition at temperatures greater than 1000 °C (which results in SO2 and O2 evolution). These results suggest organic matter is associated with the sulfate phase of these mixed salt deposits, either contained within the salt crystal structure (e.g., in fluid inclusions) or hosted on crystal surfaces/along crystal boundaries and are partially oxidised by co-evolved O2 to CO2. If associated with the salt, we propose some organic biosignatures (including FA and n-alkanes) may have been captured from the brine when salts precipitated. These biosignatures may have either nucleated or co-precipitated with the salts, becoming associated with or encapsulated within primary salt crystals, thereby protecting them against oxidation mechanisms (Keil and Mayer, 2014
Keil, R.G., Mayer, L.M. (2014) Mineral matrices and organic matter. Treatise on Geochemistry 337–359. https://doi.org/10.1016/B978-0-08-095975-7.01024-X
). The CO2 evolution in some samples could be attributed to minor carbonates, observed by LH by Battler et al. (2013)Battler, M.M., Osinski, G.R., Banerjee, N.R. (2013) Mineralogy of saline perennial cold springs on Axel Heiberg Island, Nunavut, Canada and implications for spring deposits on Mars. Icarus 224, 364–381. https://doi.org/10.1016/j.icarus.2012.08.031
. In RB-3, D-5 and D-7sub (Fig. 3b), the first CO2 peak around 650 °C could result from thermal decomposition of a Mg-bearing carbonate, and a second sharp CO2 peak around 785 °C could indicate Ca carbonate. However, in these samples there is likely organic oxidation indicated by a peak at ∼500 °C in EGA-MS traces for CO2, alkane fragments and CH4/ CH4 fragments.Figure 3 Subset representing measured samples for EGA-MS, complete sample set in Figure S-4. (a) Active Brine sample AB-1, (b) Relic Brine sample RB-3, (c) Deposit sample D-5sub, (d) D-7.1 and (e) D-7sub. All samples evolve SO2 (m/z 64), O2 (m/z 32), and CO2 (m/z 44) during heating at temperatures up to 1400 °C. All samples apart from RB-3 show the evolution of likely alkane fragments (e.g., signal at m/z 41) and possible methane (either discrete methane or CH3 fragments of a larger organic molecule; signal at m/z 15; see SI). Grey area indicates peaks of interest discussed in text.
Post-depositional recrystallisation of salts at LH may account for differences in organic profiles and EGA-MS traces observed between samples. For example, dissolution and recrystallisation can release organic compounds from close association with salt crystals at lower temperatures. The lower temperature (∼500 °C) evolution of CO2 from D-7sub (Fig. 3e), not seen in Active Brine samples, likely reflects oxidation of organics not captured within salts, which could have been released by dissolution. EGA-MS results show a single defined SO2 peak for Active Brine (Fig. 3a) samples but these peaks decrease in sharpness and definition for Relic Brine and Deposit samples (e.g., RB-3, D-7; Fig. 3b,d). Relic Brine/Deposit samples exhibit additional SO2 peaks, as exemplified by D-7sub where multiple weakly defined peaks evolve across a range of temperatures. Multiple evolutions of SO2 are suggestive of multiple sulfate phases present, which could have formed in Deposit samples as a consequence of multiple cycles of dissolution and recrystallisation in these metastable salts. Specifically, the temperatures of SO2 evolution suggest some Mg-bearing sulfates in addition to Na sulfates. Furthermore, the likely presence of carbonates in samples RB-3, D-5sub and D-7sub, which were not observed in Active Brine samples, would suggest re-working and alteration of Relic Brine and Deposit samples.
If FA are among the compounds initially trapped by salt crystallisation in Active Brine locations, such re-working would liberate them, enabling degradation and lower FA abundances observed in Relic Brine and Deposit samples. Mirabilite and thenardite, sodium sulfate minerals detected in LH salts (Fox-Powell et al., 2019)
Fox‐Powell, M.G., Osinski, G.R., Applin, D., Stromberg, J.M., Gázquez, F., et al. (2019) Natural analogue constraints on Europa’s non‐ice surface material. Geophysical Research Letters 46, 5759–5767. https://doi.org/10.1029/2018GL081339
, are sensitive to atmospheric humidity and can transition readily between hydration states. Dynamic changes to the hydration state of salt deposits over short (daily to seasonal) time scales have been observed at the LH salt deposit (Battler et al., 2013Battler, M.M., Osinski, G.R., Banerjee, N.R. (2013) Mineralogy of saline perennial cold springs on Axel Heiberg Island, Nunavut, Canada and implications for spring deposits on Mars. Icarus 224, 364–381. https://doi.org/10.1016/j.icarus.2012.08.031
; this fieldwork). The organic molecules found to survive the re-working process in the Deposit salts, distal to the active brine flow in LH salts, are n-alkanes.top
Implications for Detection of Organic Biosignatures in Planetary Salts
We have identified associations between microbial organic biosignatures and sulfate salts in a system relevant to past and present extraterrestrial brine environments. The relative abundance of FA versus n-alkanes reveals lipids evolving across active and non-active parts of the LH salt deposit. We show differences between lipids incorporated directly into precipitating salts and those remobilised by subsequent salt dissolution and recrystallisation. Our findings suggest that n-alkanes are more resilient, surviving dissolution and recrystallisation, and are found in deposits distal from active brine flow. These results have implications for the transport and preservation of organic biosignatures in low temperature planetary salts.
Encapsulated lipids in salts can record evidence of specific metabolic activities, such as microbial sulfate reduction (MSR) and autotrophic pathways, through their isotopic and molecular signatures. Additionally, coeval salt precipitation and organic capture offer insight into environmental conditions and geochemistry at the time of formation.
EGA-MS results indicate that organics are associated with sulfate salt minerals and likely co-precipitate in Active Brine samples. Salts can protect organic molecules from degradation via UV radiation and oxidising agents (Keil and Mayer, 2014
Keil, R.G., Mayer, L.M. (2014) Mineral matrices and organic matter. Treatise on Geochemistry 337–359. https://doi.org/10.1016/B978-0-08-095975-7.01024-X
). EGA-MS can also detect secondary changes in salt mineralogy, such as re-precipitation with different cations or hydration states. These changes are identified by EGA-MS through variations in thermal decomposition or thermal dehydration temperatures, as seen with Mg and Na sulfates.Studies on lipid biomarkers from hypersaline Mars analogues have also found organics preserved in sulfate salts, even in extreme acidic conditions like the Dallol hydrothermal system (Carrizo et al., 2019
Carrizo, D., Sánchez-García, L., Rodriguez, N., Gómez, F. (2019) Lipid biomarker and carbon stable isotope survey on the Dallol hydrothermal system in Ethiopia. Astrobiology 19, 1474–1489. https://doi.org/10.1089/ast.2018.1963
; Nichols et al., 2023Nichols, F., Pontefract, A., Dion‐Kirschner, H., Masterson, A.L., Osburn, M.R. (2023) Lipid Biosignatures From SO4‐Rich Hypersaline Lakes of the Cariboo Plateau. Journal of Geophysical Research: Biogeosciences 128, e2023JG007480 https://doi.org/10.1029/2023JG007480
). A hypersaline lake study using spectroscopy showed mirabilite (NaSO4) hosting organic molecules (Gill et al., 2023Gill, K.K., Jagniecki, E., Benison, K.C., Gibson, M.E. (2023) A Mars-analog sulfate mineral, mirabilite, preserves biosignatures. Geology 51, 818–822. https://doi.org/10.1130/G51256.1
). A common factor in all studies is that extreme conditions (temperature, pH, salinity) serve as strategies for lipid biomarker preservation (Finkel et al., 2023Finkel, P.L., Carrizo, D., Parro, V., Sánchez-García, L. (2023) An Overview of Lipid Biomarkers in Terrestrial Extreme Environments with Relevance for Mars Exploration. Astrobiology 23, 563–604. https://doi.org/10.1089/ast.2022.0083
).In salt deposits resulting from upwelling of subsurface brines, the transport, alteration, and distribution of molecular biosignatures will be influenced by salt crystallisation dynamics. The encapsulation of lipids within sulfate minerals implies that lipids should be most prevalent where sulfate minerals first precipitate. However, if subsequent dissolution and recrystallisation release molecules from salt crystals, there is an opportunity for post-depositional liquid-phase degradation or further transport away from the original deposition site. This is particularly important for predicting organic transport on icy worlds such as Europa, where cryovolcanic salts may have undergone fractional crystallisation or partial remobilisation of crystallised brine in the subsurface (Steinbrügge et al., 2020
Steinbrügge, G., Voigt, J.R., Wolfenbarger, N.S., Hamilton, C.W., Soderlund, K.M., et al. (2020) Brine migration and impact‐induced cryovolcanism on Europa. Geophysical Research Letters 47, e2020GL090797. https://doi.org/10.1029/2020GL090797
). Similarly, on Mars, where cycles of groundwater recharge and/or deliquescence led to dissolution and re-precipitation of evaporitic salts (Abotalib and Heggy, 2019Abotalib, A.Z., Heggy, E. (2019) A deep groundwater origin for recurring slope lineae on Mars. Nature Geoscience 12, 235–241. https://doi.org/10.1038/s41561-019-0327-5
), this could remobilise primary phase organic matter.Future missions should focus on collecting (i) samples near upwelling brines; given that FA are more prevalent where Na sulfate minerals first precipitate, (ii) salts with multiple dissolution and recrystallisation phases; to reveal long term preservation mechanisms on planetary salt deposits. Key measurements to identifying these processes will be EGA-MS, isotopic, and molecular analysis.
top
Acknowledgements
This work was funded by Leverhulme Research Project Grant RPG-2019-353. We also thank Gordon Osinski and the Polar Continental Shelf Program (Natural Resources Canada) for logistical field support.
Editor: Francis McCubbin
top
References
Abotalib, A.Z., Heggy, E. (2019) A deep groundwater origin for recurring slope lineae on Mars. Nature Geoscience 12, 235–241. https://doi.org/10.1038/s41561-019-0327-5
 Show in context
Show in context Similarly, on Mars, where cycles of groundwater recharge and/or deliquescence led to dissolution and re-precipitation of evaporitic salts (Abotalib and Heggy, 2019), this could remobilise primary phase organic matter.
View in article
Battler, M.M., Osinski, G.R., Banerjee, N.R. (2013) Mineralogy of saline perennial cold springs on Axel Heiberg Island, Nunavut, Canada and implications for spring deposits on Mars. Icarus 224, 364–381. https://doi.org/10.1016/j.icarus.2012.08.031
Show in context The CO2 evolution in some samples could be attributed to minor carbonates, observed by LH by Battler et al. (2013).
View in article
Dynamic changes to the hydration state of salt deposits over short (daily to seasonal) time scales have been observed at the LH salt deposit (Battler et al., 2013; this fieldwork).
View in article
Bibring, J.P., Langevin, Y., Mustard, J.F., Poulet, F., Arvidson, R., et al. (2006) Global mineralogical and aqueous Mars history derived from OMEGA/Mars Express data. Science 312, 400–404. https://doi.org/10.1126/science.1122659
Show in context On Mars, sulfate salts are abundant globally and record major shifts in aqueous environment chemistry (Bibring et al., 2006).
View in article
Brown, M.E., Hand, K.P. (2013) Salts and radiation products on the surface of Europa. The Astronomical Journal 145, 110. https://doi.org/10.1088/0004-6256/145/4/110
Show in context Hydrated chloride and sulfate salts are suggested to be present on the surface of Europa (Brown and Hand, 2013), originating from a range of possible endogenous and/or exogenous processes, including emplacement from subsurface liquid reservoirs (King et al., 2022).
View in article
Carrizo, D., Sánchez-García, L., Rodriguez, N., Gómez, F. (2019) Lipid biomarker and carbon stable isotope survey on the Dallol hydrothermal system in Ethiopia. Astrobiology 19, 1474–1489. https://doi.org/10.1089/ast.2018.1963
Show in context Organic biomarker results for FA and n-alkanes. Total Lipid Extract (TLE). Average Chain Length (ACL) for n-fatty acids and n-alkanes calculated following equations from Carrizo et al. (2019).
View in article
Studies on lipid biomarkers from hypersaline Mars analogues have also found organics preserved in sulfate salts, even in extreme acidic conditions like the Dallol hydrothermal system (Carrizo et al., 2019; Nichols et al., 2023).
View in article
Castañeda, I.S., Schouten, S. (2011) A review of molecular organic proxies for examining modern and ancient lacustrine environments. Quaternary Science Reviews 30, 2851–2891. https://doi.org/10.1016/j.quascirev.2011.07.009.
Show in context The general prevalence of Medium Molecular Weight (MMW; C21 to C26) over High Molecular Weight (HMW; >C27) and Low Molecular Weight (LMW; <C20) n-alkanes (Table 1) is indicative of algal and/or cyanobacteria sources (Castañeda and Shouten (2011), and references therein).
View in article
General odd/even dominance of HMW n-alkanes pointed to the contribution of higher plants and/or lichens which predominantely produce HMW n-alkanes (Castañeda and Shouten, 2011).
View in article
The LMW n-alkane distribution for all samples had no odd/even carbon number preference, suggesting derivation from microbial lipids, and/or microbial re-working of plant n-alkanes through early diagenesis (Castañeda and Shouten, 2011).
View in article
The HMW n-alkanes δ13C also suggested an input from C3 plants which usually exhibit a δ13C of −34 to −35 ‰, (Castañeda and Shouten, 2011).
View in article
Chan, Q.H., Zolensky, M.E., Kebukawa, Y., Fries, M., Ito, M., et al. (2018) Organic matter in extraterrestrial water-bearing salt crystals. Science Advances 4, eaao3521. https://doi.org/10.1126/sciadv.aao3521
Show in context Across all these targets, salt minerals have the potential to capture evidence of putative past or present microbial activity, in addition to abiogenic organic matter (Chan et al., 2018).
View in article
De Sanctis, M.C., Ammannito, E., Raponi, A., Frigeri, A., Ferrari, M., et al. (2020) Fresh emplacement of hydrated sodium chloride on Ceres from ascending salty fluids. Nature Astronomy 4, 786–793. https://doi.org/10.1038/s41550-020-1138-8
Show in context Chloride and carbonate salts sourced from subsurface water-rock reactions are components of ice grains in Enceladus plumes (Postberg et al., 2009), and carbonate and chloride salts are present on dwarf planet Ceres, likely sourced from a liquid layer beneath the crust (De Sanctis et al., 2020).
View in article
Eglinton, G., Hamilton, R.J. (1967) Leaf Epicuticular Waxes: The waxy outer surfaces of most plants display a wide diversity of fine structure and chemical constituents. Science 156, 1322–1335. https://doi.org/10.wa1126/science.156.3780.1322
Show in context Likewise samples RB-3, RB-4 and D-8 present n-alkane Cmax peaks typical of higher plants as HMW n-alkanes are known to be major components of waxes in vascular land plants (Eglinton and Hamilton, 1967).
View in article
Eigenbrode, J.L., Summons, R.E., Steele, A., Freissinet, C., Millan, M., et al. (2018) Organic matter preserved in 3-billion-year-old mudstones at Gale crater, Mars. Science 360, 1096–1101. https://doi.org/10.1126/science.aas9185
Show in context Evolved gas analysis mass spectrometry (EGA-MS) studies of salt-bearing planetary materials are currently being carried out by the Sample Analysis at Mars (SAM) instrument suite on the Mars Science Laboratory Curiosity rover (e.g., Eigenbrode et al., 2018).
View in article
Finkel, P.L., Carrizo, D., Parro, V., Sánchez-García, L. (2023) An Overview of Lipid Biomarkers in Terrestrial Extreme Environments with Relevance for Mars Exploration. Astrobiology 23, 563–604. https://doi.org/10.1089/ast.2022.0083
Show in context A common factor in all studies is that extreme conditions (temperature, pH, salinity) serve as strategies for lipid biomarker preservation (Finkel et al., 2023).
View in article
Fox‐Powell, M.G., Osinski, G.R., Applin, D., Stromberg, J.M., Gázquez, F., et al. (2019) Natural analogue constraints on Europa’s non‐ice surface material. Geophysical Research Letters 46, 5759–5767. https://doi.org/10.1029/2018GL081339
Show in context At time of sampling, the vent dome and apron deposit comprised sodium sulfates and chlorides of varying hydration states, with the highest abundances of sulfates closest to the vent (Fox-Powell et al., 2019).
View in article
Between 15 to 40 g of salt were collected for each sample in polyethylene bags from the LH deposit and maintained at 4 °C as described in Fox-Powell et al. (2019).
View in article
Mirabilite and thenardite, sodium sulfate minerals detected in LH salts (Fox-Powell et al., 2019), are sensitive to atmospheric humidity and can transition readily between hydration states.
View in article
Gill, K.K., Jagniecki, E., Benison, K.C., Gibson, M.E. (2023) A Mars-analog sulfate mineral, mirabilite, preserves biosignatures. Geology 51, 818–822. https://doi.org/10.1130/G51256.1
Show in context A hypersaline lake study using spectroscopy showed mirabilite (NaSO4) hosting organic molecules (Gill et al., 2023).
View in article
Jahnke, L.L., Des Marais, D.J. (2019) Carbon isotopic composition of lipid biomarkers from an endoevaporitic gypsum crust microbial mat reveals cycling of mineralized organic carbon. Geobiology 17, 643–659. https://doi.org/10.1111/gbi.12355
Show in context Organic biosignatures, such as lipids, are fundamental components of cell structures and are utilised as molecular markers to identify microbial activity, environmental processes, and their stable carbon isotopic ratios can reveal autotrophic carbon fixation pathways (Jahnke et al., 2019).
View in article
Furthermore, the C isotope results from LH FA (C16 and C18 from −15 to −28 ‰ and >C20 from –21 to −34 ‰) (Fig. 2 c,d), fall in the ranges for autotrophic pathways with a mixed use of three assimilation pathways of carbon including: Reverse tricarboxylic acid (rTCA) cycle for autotrophic bacteria (from −12 to −21 ‰), Calvin cycle for cyanobacteria and algae (from –19 to −30 ‰) and reductive acetyl-CoA pathway for anaerobic bacteria and archaea (−28 to −44 ‰) (Jahnke et al., 2019).
View in article
Keil, R.G., Mayer, L.M. (2014) Mineral matrices and organic matter. Treatise on Geochemistry 337–359. https://doi.org/10.1016/B978-0-08-095975-7.01024-X
Show in context These biosignatures may have either nucleated or co-precipitated with the salts, becoming associated with or encapsulated within primary salt crystals, thereby protecting them against oxidation mechanisms (Keil and Mayer, 2014).
View in article
Salts can protect organic molecules from degradation via UV radiation and oxidising agents (Keil and Mayer, 2014).
View in article
King, O., Fletcher, L.N., Ligier, N. (2022) Compositional mapping of Europa using MCMC modeling of near-IR VLT/SPHERE and Galileo/NIMS observations. The Planetary Science Journal 3, 72. https://doi.org/10.3847/PSJ/ac596d
Show in context Hydrated chloride and sulfate salts are suggested to be present on the surface of Europa (Brown and Hand, 2013), originating from a range of possible endogenous and/or exogenous processes, including emplacement from subsurface liquid reservoirs (King et al., 2022).
View in article
Lay, C.Y., Mykytczuk, N.C., Yergeau, E., Lamarche-Gagnon, G., Greer, C.W., Whyte, L.G. (2013) Defining the functional potential and active community members of a sediment microbial community in a high-arctic hypersaline subzero spring. Applied and Environmental Microbiology 79, 3637–3648. https://doi.org/10.1128/AEM.00153-13
Show in context Active microbial communities in LH vent and outflow stream are characterised by low concentrations of biomass in the sediments (around 105 cells/g) and lithotrophic microorganisms adapted to hypersaline and cold conditions: anaerobic methane oxidising archaea and sulfate reducing/sulfide oxidising bacteria are prevalent (Lay et al., 2013; Lamarche-Gagnon et al., 2015; Sapers et al., 2017; Magnuson et al., 2022).
View in article
Lamarche-Gagnon, G., Comery, R., Greer, C.W., Whyte, L.G. (2015) Evidence of in situ microbial activity and sulphidogenesis in perennially sub-0°C and hypersaline sediments of a high Arctic permafrost spring. Extremophiles 19, 1–15. https://doi.org/10.1007/s00792-014-0703-4
Show in context Active microbial communities in LH vent and outflow stream are characterised by low concentrations of biomass in the sediments (around 105 cells/g) and lithotrophic microorganisms adapted to hypersaline and cold conditions: anaerobic methane oxidising archaea and sulfate reducing/sulfide oxidising bacteria are prevalent (Lay et al., 2013; Lamarche-Gagnon et al., 2015; Sapers et al., 2017; Magnuson et al., 2022).
View in article
López-Lara, I.M., Geiger, O. (2010) Formation of fatty acids. In: Timmis, K.N. (Ed.) Handbook of Hydrocarbon and Lipid Microbiology. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-540-77587-4_26
Show in context Sources of fatty acids and n-alkanes. Fatty acids (FA) exhibited a biological signature throughout the LH salt deposit (Fig. 2a,c). This was evidenced by the presence of short-to-mid chain FA (C12-C24) of even-over-odd preference, and Cmax at C16 and C18, the dominant FA lengths for bacteria and eukaryotic cells (López-Lara and Geiger, 2010).
View in article
Magnuson, E., Altshuler, I., Fernández-Martínez, M.Á., Chen, Y.J., Maggiori, C., et al. (2022) Active lithoautotrophic and methane-oxidizing microbial community in an anoxic, sub-zero, and hypersaline High Arctic spring. The ISME Journal 16, 1798–1808. https://doi.org/10.1038/s41396-022-01233-8
Show in context Active microbial communities in LH vent and outflow stream are characterised by low concentrations of biomass in the sediments (around 105 cells/g) and lithotrophic microorganisms adapted to hypersaline and cold conditions: anaerobic methane oxidising archaea and sulfate reducing/sulfide oxidising bacteria are prevalent (Lay et al., 2013; Lamarche-Gagnon et al., 2015; Sapers et al., 2017; Magnuson et al., 2022).
View in article
The presence of i-C17, a-C17 and C18:1 at the LH Deposit corroborates past studies of sulfur isotopes and microbial analyses which identified microbial sulfate reduction and sulfur oxidation occurring in the streams and vent sediments (Moreras-Marti et al, 2021; Magnuson et al., 2022).
View in article
Moreras-Marti, A., Fox-Powell, M., Stueeken, E., Di Rocco, T., Galloway, T., et al. (2021) Quadruple sulfur isotope biosignatures from terrestrial Mars analogue systems. Geochimica et Cosmochimica Acta 308, 157–172. https://doi.org/10.1016/j.gca.2021.06.007
Show in context Quadruple Sulfur Isotope (δ34S, Δ33S and Δ36S) analysis of iron sulfide from the outflow brine support these observations with fractionations typical of microbial sulfate reduction and sulfur disproportionation (Moreras-Marti et al., 2021).
View in article
The presence of i-C17, a-C17 and C18:1 at the LH Deposit corroborates past studies of sulfur isotopes and microbial analyses which identified microbial sulfate reduction and sulfur oxidation occurring in the streams and vent sediments (Moreras-Marti et al, 2021; Magnuson et al., 2022).
View in article
Nichols, F., Pontefract, A., Dion‐Kirschner, H., Masterson, A.L., Osburn, M.R. (2023) Lipid Biosignatures From SO4‐Rich Hypersaline Lakes of the Cariboo Plateau. Journal of Geophysical Research: Biogeosciences 128, e2023JG007480 https://doi.org/10.1029/2023JG007480
Show in context Such branched FA have been identified in hypersaline lakes (Nichols et al., 2023).
View in article
Studies on lipid biomarkers from hypersaline Mars analogues have also found organics preserved in sulfate salts, even in extreme acidic conditions like the Dallol hydrothermal system (Carrizo et al., 2019; Nichols et al., 2023).
View in article
Perry, G.J., Volkman, J.K., Johns, R.B. (1979) Fatty Acids of Bacterial Origin in Contemporary Marine Sediments. Geochimica et Cosmochimica Acta 43, 1715–1725. https://doi.org/10.1016/0016-7037(79)90020-6
Show in context The i-C15, a-C15, i-C17 and a-C17 are interpreted to be associated with microbial sulfate reducers (MSR), while C18:1 records the presence of cyanobacteria or other types of bacteria and algae (Perry et al., 1979).
View in article
The rapid degradation of FA through biological processes, occurring as quickly as a few weeks, underscores their presence in terrestrial environments as potential indicators of recent microbial activity (Perry et al., 1979), such as observed in Active Brine samples, and to a lesser extent in Relic Brine samples.
View in article
Phillips, M.S., McInenly, M., Hofmann, M.H., Hinman, N.W., Warren-Rhodes, K., et al. (2023) Salt Constructs in Paleo-Lake Basins as High-Priority Astrobiology Targets. Remote Sensing 15, 314. https://doi.org/10.3390/rs15020314
Show in context Their association with liquid water environments make salts high priority astrobiology targets (Phillips et al., 2023).
View in article
Pollard, W., Omelon, C., Andersen, D., McKay, C. (1999) Perennial spring occurrence in the Expedition Fiord area of western Axel Heiberg Island, Canadian high Arctic. Canadian Journal of Earth Sciences 36, 105–120. https://doi.org/10.1139/e98-097
Show in context Axel Heiberg Island (Canadian High Arctic) hosts perennially cold hypersaline springs linked to sub-permafrost evaporite diapirs (Pollard et al., 1999).
View in article
The Lost Hammer (LH) spring (Fig. 1a) possesses the lowest temperature (−5 °C) and highest concentration of salt (24 %) of these springs, and releases gases dominated by methane (50 %) (Pollard et al., 1999).
View in article
Postberg, F., Kempf, S., Schmidt, J., Brilliantov, N., Beinsen, A., et al. (2009) Sodium salts in E-ring ice grains from an ocean below the surface of Enceladus. Nature 459, 1098–1101. https://doi.org/10.1038/nature08046
Show in context Chloride and carbonate salts sourced from subsurface water-rock reactions are components of ice grains in Enceladus plumes (Postberg et al., 2009), and carbonate and chloride salts are present on dwarf planet Ceres, likely sourced from a liquid layer beneath the crust (De Sanctis et al., 2020).
View in article
Reinhardt, M., Goetz, W., Thiel, V. (2020) Testing flight-like pyrolysis gas chromatography–mass spectrometry as performed by the Mars Organic Molecule Analyzer onboard the ExoMars 2020 rover on Oxia Planum analog samples. Astrobiology 20, 415–428. https://doi.org/10.1089/ast.2019.2143
Show in context Similar thermal analyses are planned for future in situ planetary missions (Reinhardt et al., 2020).
View in article
Sapers, H.M., Ronholm, J., Raymond-Bouchard, I., Comrey, R., Osinski, G.R., Whyte, L.G. (2017) Biological characterization of microenvironments in a hypersaline cold spring Mars analog. Frontiers in Microbiology 8, 2527.https://doi.org/10.3389/fmicb.2017.02527
Show in context Active microbial communities in LH vent and outflow stream are characterised by low concentrations of biomass in the sediments (around 105 cells/g) and lithotrophic microorganisms adapted to hypersaline and cold conditions: anaerobic methane oxidising archaea and sulfate reducing/sulfide oxidising bacteria are prevalent (Lay et al., 2013; Lamarche-Gagnon et al., 2015; Sapers et al., 2017; Magnuson et al., 2022).
View in article
Schopf, J.W., Farmer, J.D., Foster, I.S., Kudryavtsev, A.B., Gallardo, V.A., Espinoza, C. (2012) Gypsum-permineralized microfossils and their relevance to the search for life on Mars. Astrobiology 12, 619–633. https://doi.org/10.1089/ast.2012.0827
Show in context Salt minerals precipitated during evaporation or freezing of brines can capture organic and geochemical biosignatures, preserving crucial information about the aqueous environment at the time of their formation (Schopf et al., 2012).
View in article
Steinbrügge, G., Voigt, J.R., Wolfenbarger, N.S., Hamilton, C.W., Soderlund, K.M., et al. (2020) Brine migration and impact‐induced cryovolcanism on Europa. Geophysical Research Letters 47, e2020GL090797. https://doi.org/10.1029/2020GL090797
Show in context This is particularly important for predicting organic transport on icy worlds such as Europa, where cryovolcanic salts may have undergone fractional crystallisation or partial remobilisation of crystallised brine in the subsurface (Steinbrügge et al., 2020).
View in article
top
Supplementary Information
The Supplementary Information includes:
- Methodology
- Figures S-1 to S-4
- Supplementary Tables S-1 to S-3
- Supplementary Information References
Download the Supplementary Information (PDF)
Download Tables S-1 to S-3 (.xlsx)
Figures
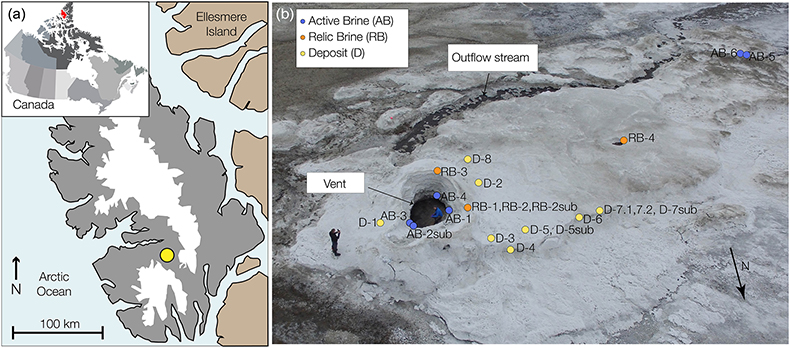
Figure 1 (a) Lost Hammer salt deposit location (yellow dot), Axel Heiberg Island. (b) Lost Hammer overview with geologist for scale. Sampling points are marked by coloured dots indicating the 3 different areas into which the Lost Hammer locality was divided: Active Brine, Relic Brine, and Deposit. Sample names ending in –sub indicate locations where a subsurface sample was paired with a corresponding surface salt.
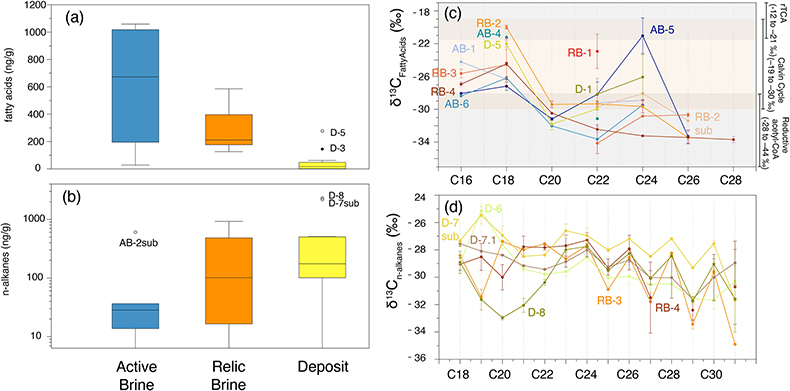
Figure 2 Whisker plot for (a) total FA and (b) total n-alkanes, concentrations in ng/g. The dots are outlier samples. Individual distributions can be found in Figures S-1, S-2 and S-3. Carbon isotopes from n-fatty acid compounds (c) and n-alkanes (d). The δ13C values for FA range between –21 ± 2.17 ‰ to –34 ± 0.01 ‰. The δ13C n-alkanes range between –25 ± 0.4 ‰ and –34 ± 0.01 ‰; complete data set in Table S-3.
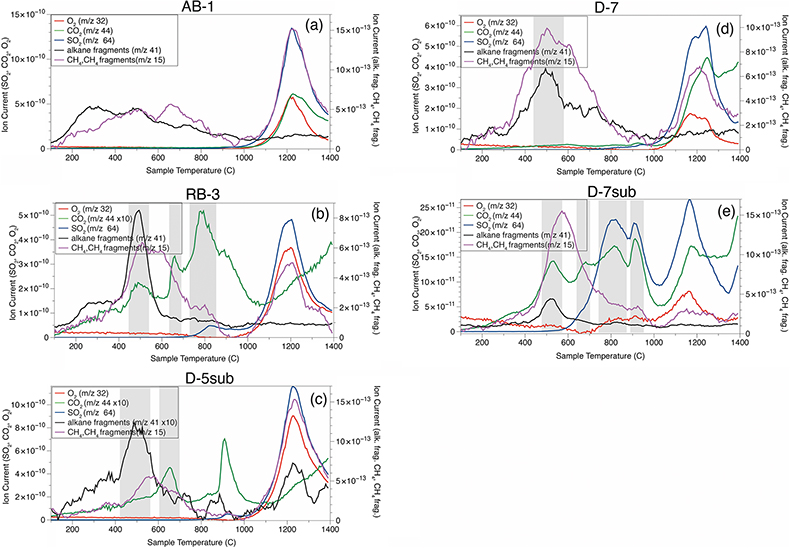
Figure 3 Subset representing measured samples for EGA-MS, complete sample set in Figure S-4. (a) Active Brine sample AB-1, (b) Relic Brine sample RB-3, (c) Deposit sample D-5sub, (d) D-7.1 and (e) D-7sub. All samples evolve SO2 (m/z 64), O2 (m/z 32), and CO2 (m/z 44) during heating at temperatures up to 1400 °C. All samples apart from RB-3 show the evolution of likely alkane fragments (e.g., signal at m/z 41) and possible methane (either discrete methane or CH3 fragments of a larger organic molecule; signal at m/z 15; see SI). Grey area indicates peaks of interest discussed in text.